Introduction
The differential susceptibility hypothesis proposed by Jay Belsky is another interpretation of psychological findings that are usually discussed according to the diathesis-stress model.
Both models suggest that people’s development and emotional affect are differentially susceptible to experiences or qualities of the environment. Where the Diathesis-stress model suggests a distinct and mostly negativity-sensitive group, Belsky describes a group that is sensitive to negative experiences but also to positive experiences. These models may be complementary, if some individuals are dually or uniquely positivity-sensitive, while other people are uniquely negativity-sensitive.
Differential Susceptibility versus Diathesis-Stress
The idea that individuals vary in their responsivity to negative qualities of the environment is generally framed in diathesis-stress or dual-risk terms. That is, some individuals, due to their biological, temperamental and/or behavioural characteristics (i.e. “diathesis” or “risk 1”), are more vulnerable to the adverse effects of negative experiences (i.e., “stress” or “risk 2”), whereas others are relatively resilient with respect to them (see Figure 1, an adaptation of Bakermans-Kranenburg and van IJzendoorn’s (2007) Figure 1). A fundamentally different, even if not competing view, of the very same phenomenon is central to Belsky’s differential susceptibility hypothesis and Boyce and Ellis’ (2005) related notion of biological sensitivity to context: Individuals do not simply vary in the degree to which they are vulnerable to the negative effects of adverse experience but, more generally, in their developmental plasticity.
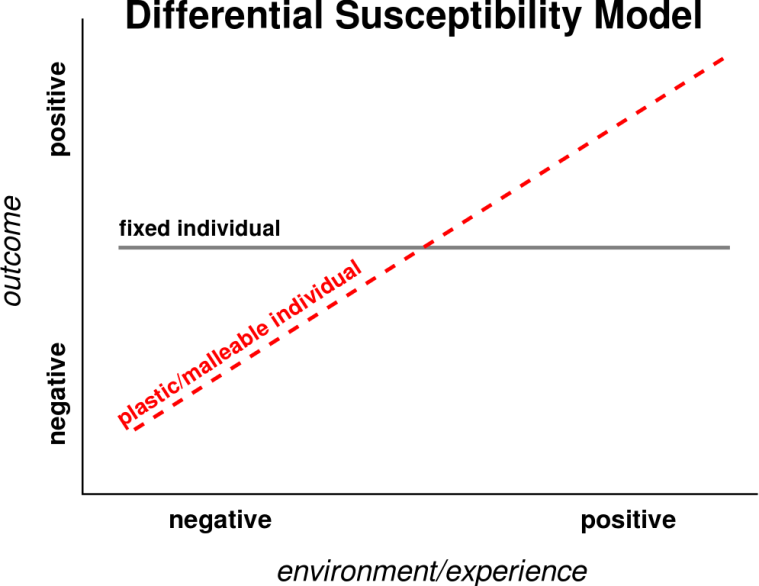
On this hypothesis, more “plastic” or malleable individuals are more susceptible than others to environmental influences in a for-better-and-for-worse manner. That is, susceptible to both the adverse developmental sequelae associated with negative environments and the positive developmental consequences of supportive ones. Less susceptible individuals, in contrast, are less affected by rearing conditions, be they presumptively supportive or undermining of well being (see Figure 2, an adaptation of Bakermans-Kranenburg and Van IJzendoorn’s (2007) Figure 1).

Theoretical Background
Belsky suggests that evolution might select for some children who are more plastic, and others who are more fixed in the face of, for example, parenting styles.
Belsky offers that ancestral parents, just like parents today, could not have known (consciously or unconsciously) which childrearing practices would prove most successful in promoting the reproductive fitness of offspring – and thus their own inclusive fitness. As a result, and as a fitness optimising strategy involving bet hedging, natural selection might have shaped parents to bear children varying in plasticity. This way, if an effect of parenting had proven counterproductive in fitness terms, those children not affected by parenting would not have incurred the cost of developing in ways that ultimately proved “misguided”.
Importantly, natural selection might favour genetic lines with both plastic and fixed developmental and affective patterns. In other words, there is value to having both kinds at once. In light of inclusive-fitness considerations, children who were less malleable (and more fixed) would have “resistance” to parental influence. This could be adaptable some times, and maladaptive other times. Their fixedness would not only have benefited themselves directly, but even their more malleable siblings indirectly. This is because siblings, like parents and children, have 50% of their genes in common. By the same token, had parenting influenced children in ways that enhanced fitness, then not only would more plastic offspring have benefited directly by following parental leads, but so, too, would their parents and even their less malleable siblings who did not benefit from the parenting they received, again for inclusive-fitness reasons. The overall effect may be to temper some of the variability in parenting. That is, to make more conservative bets.
This line of evolutionary argument leads to the prediction that children should vary in their susceptibility to parental rearing and perhaps to environmental influences more generally. As it turns out, a long line of developmental inquiry, informed by a “transactional” perspective, has more or less been based on this unstated assumption.
Criteria for the Testing of Differential Susceptibility
Belsky, Bakermans-Kranenburg, & Van IJzendoorn, (2007) delineated a series of empirical requirements – or steps – for evidencing the differential susceptibility hypothesis. Particularly they identify tests that distinguish differential susceptibility from other interaction effects including diathesis-stress/dual-risk.
While diathesis-stress/dual-risk arises when the most vulnerable are disproportionately affected in an adverse manner by a negative environment but do not also benefit disproportionately from positive environmental conditions, differential susceptibility is characterised by a cross-over interaction: the susceptible individuals are disproportionately affected by both negative and positive experiences. A further criterion that needs to be fulfilled to distinguish differential susceptibility from diathesis-stress/dual-risk is the independence of the outcome measure from the susceptibility factor: if the susceptibility factor and the outcome are related, diathesis-stress/dual-risk is suggested rather than differential susceptibility. Further, environment and susceptibility factor must also be unrelated to exclude the alternative explanation that susceptibility merely represents a function of the environment. The specificity of the differential-susceptibility effect is demonstrated if the model is not replicated when other susceptibility factors (i.e. moderators) and outcomes are used. Finally, the slope for the susceptible subgroup should be significantly different from zero and at the same time significantly steeper than the slope for the non- (or less-) susceptible subgroup.
Susceptibility Markers and Empirical Evidence
Characteristics of individuals that have been shown to moderate environmental effects in a manner consistent with the differential susceptibility hypothesis can be subdivided into three categories:
- Genetic factors;
- Endophenotypic factors; and
- Phenotypic factors.
Bakermans-Kranenburg and Van IJzendoorn (2006) were the first to test the differential susceptibility hypothesis as a function of Genetic Factors regarding the moderating effect of the dopamine receptor D4 7-repeat polymorphism (DRD4-7R) on the association between maternal sensitivity and externalising behaviour problems in 47 families. Children with the DRD4-7R allele and insensitive mothers displayed significantly more externalising behaviours than children with the same allele but with sensitive mothers. Children with the DRD4-7R allele and sensitive mothers had the least externalising behaviours of all whereas maternal sensitivity had no effect on children without the DRD4-7R allele.
Endophenotypic Factors have been examined by Obradovic, Bush, Stamperdahl, Adler and Boyce’s (2010). They investigated associations between childhood adversity and child adjustment in 338 5-year-olds. Children with high cortisol reactivity were rated by teachers as least prosocial when living under adverse conditions, but most prosocial when living under more benign conditions (and in comparison to children scoring low on cortisol reactivity).
Regarding characteristics of the category of Phenotypic Factors, Pluess and Belsky (2009) reported that the effect of child care quality on teacher-rated socioemotional adjustment varied as a function of infant temperament in the case of 761 4.5-year-olds participating in the NICHD Study of Early Child Care and Youth Development (NICHD Early Child Care Research Network, 2005). Children with difficult temperaments as infants manifest the most and least behaviour problems depending on whether they experienced, respectively, poor or good quality care (and in comparison to children with easier temperaments).


2 thoughts on “What is the Differential Susceptibility Hypothesis?”