C. Sue Carter is an American biologist and behavioural neurobiologist. She is an internationally recognised expert in behavioural neuroendocrinology. In 2014 she was appointed Director of The Kinsey Institute and Rudy Professor of Biology at Indiana University. Carter was the first person to identify the physiological mechanisms responsible for social monogamy.
Background
Carter studied biology at Drury College in Springfield, Missouri. She completed a PhD in Zoology at the University of Arkansas in Fayetteville.
Carter is a Fellow of the American Association for the Advancement of Science.
She is married to Stephen Porges, and has two children: Eric Carter Porges (currently a graduate student at the University of Chicago in Integrative Neuroscience) in Jean Decety’s Social Cognitive Neuroscience Laboratory, and Seth Porges (currently an editor at Maxim magazine in New York City, and previously an editor at Popular Mechanics magazine).
Academic Achievements
Carter studies social bonding, male and female parental behaviour, the social control of stress reactivity and the social control of reproduction, often using animal models such as the socially monogamous prairie vole. Carter’s research focuses on neuropeptide and steroid hormones, including oxytocin, vasopressin, corticotropin-releasing hormone, and oestrogen. Her research program has discovered important new developmental functions for oxytocin and vasopressin, and implicated these hormones in the regulation of long-lasting neural and effects of early social experiences. She also has a long-standing concern regarding the consequences of medical manipulations for human development and parent-child interactions, including the use of “pitocin” – a synthetic version of oxytocin – to induce labour and consequences of breastfeeding for the mother and child.
Most recently she has been examining the role of oxytocin and vasopressin in mental disorders such as autism, schizophrenia, anxiety and depression. Carter is also known for research on the physiological basis of social behaviour, including studies that implicated oxytocin, vasopressin and hormones of the hypothalamic-pituitary-adrenal (“stress”) axis in the traits of monogamy including pair-bond formation. She pioneered the physiological study of socially monogamous mammals, including the prairie vole. In collaboration with zoologist Lowell Getz, Carter documented the occurrence of social monogamy in prairie voles. Her studies in rodents helped to lay the foundation for the studies of behavioural and developmental effects of oxytocin and vasopressin in humans which are in progress. In collaboration with psychiatrist Margaret Altemus she conducted some of the first studies documenting the importance of breastfeeding in the regulation of maternal physiology.
Honours
Carter is a Fellow and Past-President of the International Behavioural Neuroscience Society and a recipient of the Matthew J. Wayner-NNOXe Pharmaceuticals Award for distinguished lifetime contributions to behavioural neuroscience.
Criticism
Author and LGBT activist Dan Savage claimed the announcement of Carter’s appointment to Director of the Kinsey Institute was “packed with bad news for anyone interested in sex research and/or conducting sex research (particularly those conducting sex research at the Kinsey Institute)” and “Carter’s pseudo-scientific/pseudo-empathetic moralizing plays right into the hands of the kind of conservative politicians who have been trying to kill the Kinsey Institute for decades.” Savage criticised Carter’s view that, “I think human sexuality must be viewed in the context of relationships,” countering, “Not all human sexuality exists in the context of relationships. You can argue, if you’re a moralist, that human sexuality should only be expressed in the context of a relationship. But that is a moral position, not a scientific one.”
This page is based on the copyrighted Wikipedia article < https://en.wikipedia.org/wiki/C._Sue_Carter >; it is used under the Creative Commons Attribution-ShareAlike 3.0 Unported License (CC-BY-SA). You may redistribute it, verbatim or modified, providing that you comply with the terms of the CC-BY-SA.
1913 – Roger Wolcott Sperry, American neuropsychologist and neurobiologist, Nobel Prize laureate (d. 1994).
People (Deaths)
1985 – Donald O. Hebb, Canadian psychologist and academic (b. 1904).
Roger Wolcott Sperry
Roger Wolcott Sperry (20 August 1913 to 17 April 1994) was an American neuropsychologist, neurobiologist and Nobel laureate who, together with David Hunter Hubel and Torsten Nils Wiesel, won the 1981 Nobel Prize in Physiology and Medicine for his work with split-brain research.
A Review of General Psychology survey, published in 2002, ranked Sperry as the 44th most cited psychologist of the 20th century.
Donald O. Hebb
Donald Olding Hebb FRS (22 July 1904 to 20 August 1985) was a Canadian psychologist who was influential in the area of neuropsychology, where he sought to understand how the function of neurons contributed to psychological processes such as learning.
He is best known for his theory of Hebbian learning, which he introduced in his classic 1949 work The Organisation of Behaviour. He has been described as the father of neuropsychology and neural networks. A Review of General Psychology survey, published in 2002, ranked Hebb as the 19th most cited psychologist of the 20th century. His views on learning described behaviour and thought in terms of brain function, explaining cognitive processes in terms of connections between neuron assemblies.
Clinical neuroscience is a branch of neuroscience that focuses on the scientific study of fundamental mechanisms that underlie diseases and disorders of the brain and central nervous system. It seeks to develop new ways of conceptualising and diagnosing such disorders and ultimately of developing novel treatments.
Background
A clinical neuroscientist is a scientist who has specialised knowledge in the field. Not all clinicians are clinical neuroscientists. Clinicians and scientists – including psychiatrists, neurologists, clinical psychologists, neuroscientists, and other specialists – use basic research findings from neuroscience in general and clinical neuroscience in particular to develop diagnostic methods and ways to prevent and treat neurobiological disorders. Such disorders include addiction, Alzheimer’s disease, amyotrophic lateral sclerosis, anxiety disorders, attention deficit hyperactivity disorder, autism, bipolar disorder, brain tumours, depression, Down syndrome, dyslexia, epilepsy, Huntington’s disease, multiple sclerosis, neurological AIDS, neurological trauma, pain, obsessive-compulsive disorder, Parkinson’s disease, schizophrenia, sleep disorders, stroke and Tourette syndrome.
While neurology, neurosurgery and psychiatry are the main medical specialties that use neuroscientific information, other specialties such as cognitive neuroscience, neuroradiology, neuropathology, ophthalmology, otorhinolaryngology, anaesthesiology and rehabilitation medicine can contribute to the discipline. Integration of the neuroscience perspective alongside other traditions like psychotherapy, social psychiatry or social psychology will become increasingly important.
One Mind for Research
The “One Mind for Research” forum was a convention held in Boston, Massachusetts on 23 to 25 May 2011 that produced the blueprint document A Ten-Year Plan for Neuroscience: From Molecules to Brain Health. Leading neuroscience researchers and practitioners in the United States contributed to the creation of this document, in which 17 key areas of opportunities are listed under the Clinical Neuroscience section. These include the following:
Rethinking curricula to break down intellectual silos.
Training translational neuroscientists and clinical investigators.
Investigating biomarkers.
Improving psychiatric diagnosis.
Developing a “Framingham Study of Brain Disorders” (i.e. longitudinal cohort for central nervous system disease).
Identifying developmental risk factors and producing effective interventions.
Discovering new treatments for pain, including neuropathic pain.
Treating disorders of neural signalling and pathological synchrony.
Treating disorders of immunity or inflammation.
Treating metabolic and mitochondrial disorders.
Developing new treatments for depression.
Treating addictive disorders.
Improving treatment of schizophrenia.
Preventing and treating cerebrovascular disease.
Achieving personalized medicine.
Understanding shared mechanisms of neurodegeneration.
Advancing anaesthesia.
In particular, it advocates for better integrated and scientifically driven curricula for practitioners, and it recommends that such curricula be shared among neurologists, psychiatrists, psychologists, neurosurgeons and neuroradiologists.
Given the various ethical, legal and societal implications for healthcare practitioners arising from advances in neuroscience, the University of Pennsylvania inaugurated the Penn Conference on Clinical Neuroscience and Society in July 2011.
This page is based on the copyrighted Wikipedia article <https://en.wikipedia.org/wiki/Clinical_neuroscience >; it is used under the Creative Commons Attribution-ShareAlike 3.0 Unported License (CC-BY-SA). You may redistribute it, verbatim or modified, providing that you comply with the terms of the CC-BY-SA.
Scientific studies have found that different brain areas show altered activity in people with major depressive disorder (MDD), and this has encouraged advocates of various theories that seek to identify a biochemical origin of the disease, as opposed to theories that emphasize psychological or situational causes.
Factors spanning these causative groups include nutritional deficiencies in magnesium, vitamin D, and tryptophan with situational origin but biological impact. Several theories concerning the biologically based cause of depression have been suggested over the years, including theories revolving around monoamine neurotransmitters, neuroplasticity, neurogenesis, inflammation and the circadian rhythm. Physical illnesses, including hypothyroidism and mitochondrial disease, can also trigger depressive symptoms.
Neural circuits implicated in depression include those involved in the generation and regulation of emotion, as well as in reward. Abnormalities are commonly found in the lateral prefrontal cortex whose putative function is generally considered to involve regulation of emotion. Regions involved in the generation of emotion and reward such as the amygdala, anterior cingulate cortex (ACC), orbitofrontal cortex (OFC), and striatum are frequently implicated as well. These regions are innervated by a monoaminergic nuclei, and tentative evidence suggests a potential role for abnormal monoaminergic activity.
Genetic Factors
Difficulty of Gene Studies
Historically, candidate gene studies have been a major focus of study. However, as the number of genes reduces the likelihood of choosing a correct candidate gene, Type I errors (false positives) are highly likely. Candidate genes studies frequently possess a number of flaws, including frequent genotyping errors and being statistically underpowered. These effects are compounded by the usual assessment of genes without regard for gene-gene interactions. These limitations are reflected in the fact that no candidate gene has reached genome-wide significance.
Gene Candidates
5-HTTLPR
The 5-HTTLPR, or serotonin transporter promoter gene’s short allele, has been associated with increased risk of depression; since the 1990s, however, results have been inconsistent. Other genes that have been linked to a gene-environment interaction include CRHR1, FKBP5 and BDNF, the first two of which are related to the stress reaction of the HPA axis, and the latter of which is involved in neurogenesis. Candidate gene analysis of 5-HTTLPR on depression was inconclusive on its effect, either alone or in combination with life stress.
A 2003 study proposed that a gene-environment interaction (GxE) may explain why life stress is a predictor for depressive episodes in some individuals, but not in others, depending on an allelic variation of the serotonin-transporter-linked promoter region (5-HTTLPR). This hypothesis was widely-discussed in both the scientific literature and popular media, where it was dubbed the “Orchid gene”, but has conclusively failed to replicate in much larger samples, and the observed effect sizes in earlier work are not consistent with the observed polygenicity of depression.
BDNF
BDNF polymorphisms have also been hypothesized to have a genetic influence, but early findings and research failed to replicate in larger samples, and the effect sizes found by earlier estimates are inconsistent with the observed polygenicity of depression.
SIRT1 and LHPP
A 2015 GWAS study in Han Chinese women positively identified two variants in intronic regions near SIRT1 and LHPP with a genome-wide significant association.
Norepinephrine Transporter Polymorphisms
Attempts to find a correlation between norepinephrine transporter polymorphisms and depression have yielded negative results.
One review identified multiple frequently studied candidate genes. The genes encoding for the 5-HTT and 5-HT2A receptor were inconsistently associated with depression and treatment response. Mixed results were found for brain-derived neurotrophic factor (BDNF) Val66Met polymorphisms. Polymorphisms in the tryptophan hydroxylase gene was found to be tentatively associated with suicidal behaviour. A meta analysis of 182 case controlled genetic studies published in 2008 found Apolipoprotein E verepsilon 2 to be protective, and GNB3 825T, MTHFR 677T, SLC6A4 44bp insertion or deletions, and SLC6A3 40 bpVNTR 9/10 genotype to confer risk.
Circadian Rhythm
Depression may be related to abnormalities in the circadian rhythm, or biological clock.
A well synchronised circadian rhythm is critical for maintaining optimal health. Adverse changes and alterations in the circadian rhythm have been associated various neurological disorders and mood disorders including depression.
Depression may be related to the same brain mechanisms that control the cycles of sleep and wakefulness.
Sleep
Sleep disturbance is the most prominent symptom in depressive patients. Studies about sleep electroencephalograms have shown characteristic changes in depression such as reductions in non-rapid eye movement sleep production, disruptions of sleep continuity and disinhibition of rapid eye movement (REM) sleep. Rapid eye movement (REM) sleep – the stage in which dreaming occurs – may be quick to arrive and intense in depressed people. REM sleep depends on decreased serotonin levels in the brain stem, and is impaired by compounds, such as antidepressants, that increase serotonergic tone in brain stem structures. Overall, the serotonergic system is least active during sleep and most active during wakefulness. Prolonged wakefulness due to sleep deprivation activates serotonergic neurons, leading to processes similar to the therapeutic effect of antidepressants, such as the selective serotonin reuptake inhibitors (SSRIs). Depressed individuals can exhibit a significant lift in mood after a night of sleep deprivation. SSRIs may directly depend on the increase of central serotonergic neurotransmission for their therapeutic effect, the same system that impacts cycles of sleep and wakefulness.
Light Therapy
Research on the effects of light therapy on seasonal affective disorder suggests that light deprivation is related to decreased activity in the serotonergic system and to abnormalities in the sleep cycle, particularly insomnia. Exposure to light also targets the serotonergic system, providing more support for the important role this system may play in depression. Sleep deprivation and light therapy both target the same brain neurotransmitter system and brain areas as antidepressant drugs, and are now used clinically to treat depression. Light therapy, sleep deprivation and sleep time displacement (sleep phase advance therapy) are being used in combination quickly to interrupt a deep depression in people who are hospitalised for MDD.
Increased and decreased sleep length appears to be a risk factor for depression. People with MDD sometimes show diurnal and seasonal variation of symptom severity, even in non-seasonal depression. Diurnal mood improvement was associated with activity of dorsal neural networks. Increased mean core temperature was also observed. One hypothesis proposed that depression was a result of a phase shift.
Daytime light exposure correlates with decreased serotonin transporter activity, which may underlie the seasonality of some depression.
Monoamines
Monoamines are neurotransmitters that include serotonin, dopamine, norepinephrine, and epinephrine.
Illustration of the major elements in a prototypical synapse. Synapses are gaps between nerve cells. These cells convert their electrical impulses into bursts of chemical relayers, called neurotransmitters, which travel across the synapses to receptors on adjacent cells, triggering electrical impulses to travel down the latter cells.
Monoamine Hypothesis of Depression
Many antidepressant drugs acutely increase synaptic levels of the monoamine neurotransmitter, serotonin, but they may also enhance the levels of norepinephrine and dopamine. The observation of this efficacy led to the monoamine hypothesis of depression, which postulates that the deficit of certain neurotransmitters is responsible for depression, and even that certain neurotransmitters are linked to specific symptoms. Normal serotonin levels have been linked to mood and behaviour regulation, sleep, and digestion; norepinephrine to the fight-or-flight response; and dopamine to movement, pleasure, and motivation. Some have also proposed the relationship between monoamines and phenotypes such as serotonin in sleep and suicide, norepinephrine in dysphoria, fatigue, apathy, cognitive dysfunction, and dopamine in loss of motivation and psychomotor symptoms.[31] The main limitation for the monoamine hypothesis of depression is the therapeutic lag between initiation of antidepressant treatment and perceived improvement of symptoms. One explanation for this therapeutic lag is that the initial increase in synaptic serotonin is only temporary, as firing of serotonergic neurons in the dorsal raphe adapt via the activity of 5-HT1A autoreceptors. The therapeutic effect of antidepressants is thought to arise from autoreceptor desensitization over a period of time, eventually elevating firing of serotonergic neurons.
Serotonin
Initial studies of serotonin in depression examined peripheral measures such as the serotonin metabolite 5-Hydroxyindoleacetic acid (5-HIAA) and platelet binding. The results were generally inconsistent, and may not generalise to the central nervous system. However evidence from receptor binding studies and pharmacological challenges provide some evidence for dysfunction of serotonin neurotransmission in depression. Serotonin may indirectly influence mood by altering emotional processing biases that are seen at both the cognitive/behavioural and neural level. Pharmacologically reducing serotonin synthesis, and pharmacologically enhancing synaptic serotonin can produce and attenuate negative affective biases, respectively. These emotional processing biases may explain the therapeutic gap.
Dopamine
While various abnormalities have been observed in dopaminergic systems, results have been inconsistent. People with MDD have an increased reward response to dextroamphetamine compared to controls, and it has been suggested that this results from hypersensitivity of dopaminergic pathways due to natural hypoactivity. While polymorphisms of the D4 and D3 receptor have been implicated in depression, associations have not been consistently replicated. Similar inconsistency has been found in post-mortem studies, but various dopamine receptor agonists show promise in treating MDD. There is some evidence that there is decreased nigrostriatal pathway activity in people with melancholic depression (psychomotor retardation). Further supporting the role of dopamine in depression is the consistent finding of decreased cerebrospinal fluid and jugular metabolites of dopamine, as well as post mortem findings of altered Dopamine receptor D3 and dopamine transporter expression. Studies in rodents have supported a potential mechanism involving stress-induced dysfunction of dopaminergic systems.
Monoamine receptors affect phospholipase C and adenylyl cyclase inside of the cell. Green arrows means stimulation and red arrows inhibition. Serotonin receptors are blue, norepinephrine orange, and dopamine yellow. Phospholipase C and adenylyl cyclase start a signalling cascade which turn on or off genes in the cell. Sufficient ATP from mitochondria is required for these downstream signalling events. The 5HT-3 receptor is associated with gastrointestinal adverse effects and has no relationship to the other monoamine receptors.
Catecholamines
A number of lines of evidence indicative of decreased adrenergic activity in depression have been reported. Findings include the decreased activity of tyrosine hydroxylase, decreased size of the locus coeruleus, increased alpha 2 adrenergic receptor density, and decreased alpha 1 adrenergic receptor density. Furthermore, norepinephrine transporter knockout in mice models increases their tolerance to stress, implicating norepinephrine in depression.
One method used to study the role of monoamines is monoamine depletion. Depletion of tryptophan (the precursor of serotonin), tyrosine and phenylalanine (precursors to dopamine) does result in decreased mood in those with a predisposition to depression, but not in persons lacking the predisposition. On the other hand, inhibition of dopamine and norepinephrine synthesis with alpha-methyl-para-tyrosine does not consistently result in decreased mood.
Monoamine Oxidase
An offshoot of the monoamine hypothesis suggests that monoamine oxidase A (MAO-A), an enzyme which metabolises monoamines, may be overly active in depressed people. This would, in turn, cause the lowered levels of monoamines. This hypothesis received support from a PET study, which found significantly elevated activity of MAO-A in the brain of some depressed people. In genetic studies, the alterations of MAO-A-related genes have not been consistently associated with depression. Contrary to the assumptions of the monoamine hypothesis, lowered but not heightened activity of MAO-A was associated with depressive symptoms in adolescents. This association was observed only in maltreated youth, indicating that both biological (MAO genes) and psychological (maltreatment) factors are important in the development of depressive disorders. In addition, some evidence indicates that disrupted information processing within neural networks, rather than changes in chemical balance, might underlie depression.
Limitations
Since the 1990s, research has uncovered multiple limitations of the monoamine hypothesis, and its inadequacy has been criticised within the psychiatric community. For one thing, serotonin system dysfunction cannot be the sole cause of depression. Not all patients treated with antidepressants show improvements despite the usually rapid increase in synaptic serotonin. If significant mood improvements do occur, this is often not for at least two to four weeks. One possible explanation for this lag is that the neurotransmitter activity enhancement is the result of auto receptor desensitization, which can take weeks. Intensive investigation has failed to find convincing evidence of a primary dysfunction of a specific monoamine system in people with MDD. The antidepressants that do not act through the monoamine system, such as tianeptine and opipramol, have been known for a long time. There have also been inconsistent findings with regard to levels of serum 5-HIAA, a metabolite of serotonin. Experiments with pharmacological agents that cause depletion of monoamines have shown that this depletion does not cause depression in healthy people. Another problem that presents is that drugs that deplete monoamines may actually have antidepressant properties. Further, some have argued that depression may be marked by a hyperserotonergic state. Already limited, the monoamine hypothesis has been further oversimplified when presented to the general public.
Receptor Binding
As of 2012, efforts to determine differences in neurotransmitter receptor expression or for function in the brains of people with MDD using positron emission tomography (PET) had shown inconsistent results. Using the PET imaging technology and reagents available as of 2012, it appeared that the D1 receptor may be under-expressed in the striatum of people with MDD. 5-HT1A receptor binding literature is inconsistent; however, it leans towards a general decrease in the mesiotemporal cortex. 5-HT2A receptor binding appears to be unregulated in people with MDD. Results from studies on 5-HTT binding are variable, but tend to indicate higher levels in people with MDD. Results with D2/D3 receptor binding studies are too inconsistent to draw any conclusions. Evidence supports increased MAO activity in people with MDD, and it may even be a trait marker (not changed by response to treatment). Muscarinic receptor binding appears to be increased in depression, and, given ligand binding dynamics, suggests increased cholinergic activity.
Four meta analyses on receptor binding in depression have been performed, two on serotonin transporter (5-HTT), one on 5-HT1A, and another on dopamine transporter (DAT). One meta analysis on 5-HTT reported that binding was reduced in the midbrain and amygdala, with the former correlating with greater age, and the latter correlating with depression severity. Another meta-analysis on 5-HTT including both post-mortem and in vivo receptor binding studies reported that while in vivo studies found reduced 5-HTT in the striatum, amygdala and midbrain, post mortem studies found no significant associations. 5-HT1A was found to be reduced in the anterior cingulate cortex, mesiotemporal lobe, insula, and hippocampus, but not in the amygdala or occipital lobe. The most commonly used 5-HT1A ligands are not displaced by endogenous serotonin, indicating that receptor density or affinity is reduced. Dopamine transporter binding is not changed in depression.
Emotional Processing and Neural Circuits
Emotional Bias
People with MDD show a number of biases in emotional processing, such as a tendency to rate happy faces more negatively, and a tendency to allocate more attentional resources to sad expressions. Depressed people also have impaired recognition of happy, angry, disgusted, fearful and surprised, but not sad faces. Functional neuroimaging has demonstrated hyperactivity of various brain regions in response to negative emotional stimuli, and hypoactivity in response to positive stimuli. One meta analysis reported that depressed subjects showed decreased activity in the left dorsolateral prefrontal cortex and increased activity in the amygdala in response to negative stimuli. Another meta analysis reported elevated hippocampus and thalamus activity in a subgroup of depressed subjects who were medication naïve, not elderly, and had no comorbidities. The therapeutic lag of antidepressants has been suggested to be a result of antidepressants modifying emotional processing leading to mood changes. This is supported by the observation that both acute and sub-chronic SSRI administration increases response to positive faces. Antidepressant treatment appears to reverse mood congruent biases in limbic, prefrontal, and fusiform areas. dlPFC response is enhanced and amygdala response is attenuated during processing of negative emotions, the former or which is thought to reflect increased top down regulation. The fusiform gyrus and other visual processing areas respond more strongly to positive stimuli with antidepressant treatment, which is thought to reflect the a positive processing bias. These effects do not appear to be unique to serotonergic or noradrenergic antidepressants, but also occur in other forms of treatment such as deep brain stimulation.
Neural Circuits
One meta analysis of functional neuroimaging in depression observed a pattern of abnormal neural activity hypothesized to reflect an emotional processing bias. Relative to controls, people with MDD showed hyperactivity of circuits in the salience network (SN), composed of the pulvinar nuclei, the insula, and the dorsal anterior cingulate cortex (dACC), as well as decreased activity in regulatory circuits composed of the striatum and dlPFC.
A neuroanatomical model called the limbic-cortical model has been proposed to explain early biological findings in depression. The model attempts to relate specific symptoms of depression to neurological abnormalities. Elevated resting amygdala activity was proposed to underlie rumination, as stimulation of the amygdala has been reported to be associated with the intrusive recall of negative memories. The ACC was divided into pregenual (pgACC) and subgenual regions (sgACC), with the former being electrophysiologically associated with fear, and the latter being metabolically implicated in sadness in healthy subjects. Hyperactivity of the lateral orbitofrontal and insular regions, along with abnormalities in lateral prefrontal regions was suggested to underlie maladaptive emotional responses, given the regions roles in reward learning. This model and another termed “the cortical striatal model”, which focused more on abnormalities in the cortico-basal ganglia-thalamo-cortical loop, have been supported by recent literature. Reduced striatal activity, elevated OFC activity, and elevated sgACC activity were all findings consistent with the proposed models. However, amygdala activity was reported to be decreased, contrary to the limbic-cortical model. Furthermore, only lateral prefrontal regions were modulated by treatment, indicating that prefrontal areas are state markers (i.e. dependent upon mood), while subcortical abnormalities are trait markers (i.e. reflect a susceptibility).
Reward
While depression severity as a whole is not correlated with a blunted neural response to reward, anhedonia is directly correlated to reduced activity in the reward system. The study of reward in depression is limited by heterogeneity in the definition and conceptualisations of reward and anhedonia. Anhedonia is broadly defined as a reduced ability to feel pleasure, but questionnaires and clinical assessments rarely distinguish between motivational “wanting” and consummatory “liking”. While a number of studies suggest that depressed subjects rate positive stimuli less positively and as less arousing, a number of studies fail to find a difference. Furthermore, response to natural rewards such as sucrose does not appear to be attenuated. General affective blunting may explain “anhedonic” symptoms in depression, as meta analysis of both positive and negative stimuli reveal reduced rating of intensity. As anhedonia is a prominent symptom of depression, direct comparison of depressed with healthy subjects reveals increased activation of the subgenual anterior cingulate cortex (sgACC), and reduced activation of the ventral striatum, and in particular the nucleus accumbens (NAcc) in response to positive stimuli. Although the finding of reduced NAcc activity during reward paradigms is fairly consistent, the NAcc is made up of a functionally diverse range of neurons, and reduced blood-oxygen-level dependent (BOLD) signal in this region could indicate a variety of things including reduced afferent activity or reduced inhibitory output. Nevertheless, these regions are important in reward processing, and dysfunction of them in depression is thought to underlie anhedonia. Residual anhedonia that is not well targeted by serotonergic antidepressants is hypothesized to result from inhibition of dopamine release by activation of 5-HT2C receptors in the striatum. The response to reward in the medial orbitofrontal cortex (OFC) is attenuated in depression, while lateral OFC response is enhanced to punishment. The lateral OFC shows sustained response to absence of reward or punishment, and it is thought to be necessary for modifying behaviour in response to changing contingencies. Hypersensitivity in the lOFC may lead to depression by producing a similar effect to learned helplessness in animals.
Elevated response in the sgACC is a consistent finding in neuroimaging studies using a number of paradigms including reward related tasks. Treatment is also associated with attenuated activity in the sgACC, and inhibition of neurons in the rodent homologue of the sgACC, the infralimbic cortex (IL), produces an antidepressant effect. Hyperactivity of the sgACC has been hypothesized to lead to depression via attenuating the somatic response to reward or positive stimuli. Contrary to studies of functional magnetic resonance imaging response in the sgACC during tasks, resting metabolism is reduced in the sgACC. However, this is only apparent when correcting for the prominent reduction in sgACC volume associated with depression; structural abnormalities are evident at a cellular level, as neuropathological studies report reduced sgACC cell markers. The model of depression proposed from these findings by Drevets et al. suggests that reduced sgACC activity results in enhanced sympathetic nervous system activity and blunted HPA axis feedback. Activity in the sgACC may also not be causal in depression, as the authors of one review that examined neuroimaging in depressed subjects during emotional regulation hypothesized that the pattern of elevated sgACC activity reflected increased need to modulate automatic emotional responses in depression. More extensive sgACC and general prefrontal recruitment during positive emotional processing was associated with blunted subcortical response to positive emotions, and subject anhedonia. This was interpreted by the authors to reflect a downregulation of positive emotions by the excessive recruitment of the prefrontal cortex.
Neuroanatomy
While a number of neuroimaging findings are consistently reported in people with major depressive disorder, the heterogeneity of depressed populations presents difficulties interpreting these findings. For example, averaging across populations may hide certain subgroup related findings; while reduced dlPFC activity is reported in depression, a subgroup may present with elevated dlPFC activity. Averaging may also yield statistically significant findings, such as reduced hippocampal volumes, that are actually present in a subgroup of subjects. Due to these issues and others, including the longitudinal consistency of depression, most neural models are likely inapplicable to all depression.
Structural Neuroimaging
Meta analyses performed using seed-based d mapping have reported grey matter reductions in a number of frontal regions. One meta analysis of early onset general depression reported grey matter reductions in the bilateral anterior cingulate cortex (ACC) and dorsomedial prefrontal cortex (dmPFC). One meta analysis on first episode depression observed distinct patterns of grey matter reductions in medication free, and combined populations; medication free depression was associated with reductions in the right dorsolateral prefrontal cortex, right amygdala, and right inferior temporal gyrus; analysis on a combination of medication free and medicated depression found reductions in the left insula, right supplementary motor area, and right middle temporal gyrus. Another review distinguishing medicated and medication free populations, albeit not restricted to people with their first episode of MDD, found reductions in the combined population in the bilateral superior, right middle, and left inferior frontal gyrus, along with the bilateral parahippocampus. Increases in thalamic and ACC grey matter was reported in the medication free and medicated populations respectively. A meta analysis performed using “activation likelihood estimate” reported reductions in the paracingulate cortex, dACC and amygdala.
GMV reductions in MDD and BD.
Using statistical parametric mapping, one meta analysis replicated previous findings of reduced grey matter in the ACC, medial prefrontal cortex, inferior frontal gyrus, hippocampus and thalamus; however reductions in the OFC and ventromedial prefrontal cortex grey matter were also reported.
Two studies on depression from the ENIGMA consortium have been published, one on cortical thickness, and the other on subcortical volume. Reduced cortical thickness was reported in the bilateral OFC, ACC, insula, middle temporal gyri, fusiform gyri, and posterior cingulate cortices, while surface area deficits were found in medial occipital, inferior parietal, orbitofrontal and precentral regions. Subcortical abnormalities, including reductions in hippocampus and amygdala volumes, which were especially pronounced in early onset depression.
Multiple meta analysis have been performed on studies assessing white matter integrity using fractional anisotropy (FA). Reduced FA has been reported in the corpus callosum (CC) in both first episode medication naïve, and general major depressive populations. The extent of CC reductions differs from study to study. People with MDD who have not taken antidepressants before have been reported to have reductions only in the body of the CC and only in the genu of the CC. On the other hand, general MDD samples have been reported to have reductions in the body of the CC, the body and genu of the CC, and only the genu of the CC. Reductions of FA have also been reported in the anterior limb of the internal capsule (ALIC) and superior longitudinal fasciculus.
Functional Neuroimaging
Studies of resting state activity have utilised a number of indicators of resting state activity, including regional homogeneity (ReHO), amplitude of low frequency fluctuations (ALFF), fractional amplitude of low frequency fluctuations (fALFF), arterial spin labelling (ASL), and positron emission tomography measures of regional cerebral blood flow or metabolism.
MDD is associated with reduced FA in the ALIC and genu/body of the CC.
Studies using ALFF and fALFF have reported elevations in ACC activity, with the former primarily reporting more ventral findings, and the latter more dorsal findings. A conjunction analysis of ALFF and CBF studies converged on the left insula, with previously untreated people having increased insula activity. Elevated caudate CBF was also reported A meta analysis combining multiple indicators of resting activity reported elevated anterior cingulate, striatal, and thalamic activity and reduced left insula, post-central gyrus and fusiform gyrus activity. An activation likelihood estimate (ALE) meta analysis of PET/SPECT resting state studies reported reduced activity in the left insula, pregenual and dorsal anterior cingulate cortex and elevated activity in the thalamus, caudate, anterior hippocampus and amygdala. Compared to the ALE meta analysis of PET/SPECT studies, a study using multi-kernel density analysis reported hyperactivity only in the pulvinar nuclei of the thalamus.
Brain Regions
Research on the brains of people with MDD usually shows disturbed patterns of interaction between multiple parts of the brain. Several areas of the brain are implicated in studies seeking to more fully understand the biology of depression:
Subgenual Cingulate
Studies have shown that Brodmann area 25, also known as subgenual cingulate, is metabolically overactive in treatment-resistant depression. This region is extremely rich in serotonin transporters and is considered as a governor for a vast network involving areas like hypothalamus and brain stem, which influences changes in appetite and sleep; the amygdala and insula, which affect the mood and anxiety; the hippocampus, which plays an important role in memory formation; and some parts of the frontal cortex responsible for self-esteem. Thus disturbances in this area or a smaller than normal size of this area contributes to depression. Deep brain stimulation has been targeted to this region in order to reduce its activity in people with treatment resistant depression.
Prefrontal Cortex
One review reported hypoactivity in the prefrontal cortex of those with depression compared to controls. The prefrontal cortex is involved in emotional processing and regulation, and dysfunction of this process may be involved in the aetiology of depression. One study on antidepressant treatment found an increase in PFC activity in response to administration of antidepressants. One meta analysis published in 2012 found that areas of the prefrontal cortex were hypoactive in response to negative stimuli in people with MDD. One study suggested that areas of the prefrontal cortex are part of a network of regions including dorsal and pregenual cingulate, bilateral middle frontal gyrus, insula and superior temporal gyrus that appear to be hypoactive in people with MDD. However the authors cautioned that the exclusion criteria, lack of consistency and small samples limit results.
Amygdala
The amygdala, a structure involved in emotional processing appears to be hyperactive in those with major depressive disorder. The amygdala in unmedicated depressed persons tended to be smaller than in those that were medicated, however aggregate data shows no difference between depressed and healthy persons. During emotional processing tasks right amygdala is more active than the left, however there is no differences during cognitive tasks, and at rest only the left amygdala appears to be more hyperactive. One study, however, found no difference in amygdala activity during emotional processing tasks.
Hippocampus
Atrophy of the hippocampus has been observed during depression, consistent with animal models of stress and neurogenesis.
Stress can cause depression and depression-like symptoms through monoaminergic changes in several key brain regions as well as suppression in hippocampal neurogenesis. This leads to alteration in emotion and cognition related brain regions as well as HPA axis dysfunction. Through the dysfunction, the effects of stress can be exacerbated including its effects on 5-HT. Furthermore, some of these effects are reversed by antidepressant action, which may act by increasing hippocampal neurogenesis. This leads to a restoration in HPA activity and stress reactivity, thus restoring the deleterious effects induced by stress on 5-HT.
The hypothalamic-pituitary-adrenal axis is a chain of endocrine structures that are activated during the body’s response to stressors of various sorts. The HPA axis involves three structure, the hypothalamus which release CRH that stimulates the pituitary gland to release ACTH which stimulates the adrenal glands to release cortisol. Cortisol has a negative feedback effect on the pituitary gland and hypothalamus. In people with MDD this often shows increased activation in depressed people, but the mechanism behind this is not yet known. Increased basal cortisol levels and abnormal response to dexamethasone challenges have been observed in people with MDD. Early life stress has been hypothesized as a potential cause of HPA dysfunction. HPA axis regulation may be examined through a dexamethasone suppression tests, which tests the feedback mechanisms. Non-suppression of dexamethasone is a common finding in depression, but is not consistent enough to be used as a diagnostic tool. HPA axis changes may be responsible for some of the changes such as decreased bone mineral density and increased weight found in people with MDD. One drug, ketoconazole, currently under development has shown promise in treating MDD.
Hippocampal Neurogenesis
Reduced hippocampal neurogenesis leads to a reduction in hippocampal volume. A genetically smaller hippocampus has been linked to a reduced ability to process psychological trauma and external stress, and subsequent predisposition to psychological illness. Depression without familial risk or childhood trauma has been linked to a normal hippocampal volume but localised dysfunction.
Animal Models
A number of animal models exist for depression, but they are limited in that depression involves primarily subjective emotional changes. However, some of these changes are reflected in physiology and behaviour, the latter of which is the target of many animal models. These models are generally assessed according to four facets of validity; the reflection of the core symptoms in the model; the predictive validity of the model; the validity of the model with regard to human characteristics of aetiology; and the biological plausibility.
Different models for inducing depressive behaviours have been utilised; neuroanatomical manipulations such as olfactory bulbectomy or circuit specific manipulations with optogenetics; genetic models such as 5-HT1A knockout or selectively bred animals; models involving environmental manipulation associated with depression in humans, including chronic mild stress, early life stress and learned helplessness. The validity of these models in producing depressive behaviours may be assessed with a number of behavioural tests. Anhedonia and motivational deficits may, for example, be assessed via examining an animal’s level of engagement with rewarding stimuli such as sucrose or intracranial self-stimulation. Anxious and irritable symptoms may be assessed with exploratory behaviour in the presence of a stressful or novelty environment, such as the open field test, novelty suppressed feeding, or the elevated plus-maze. Fatigue, psychomotor poverty, and agitation may be assessed with locomotor activity, grooming activity, and open field tests.
Animal models possess a number of limitations due to the nature of depression. Some core symptoms of depression, such as rumination, low self-esteem, guilt, and depressed mood cannot be assessed in animals as they require subjective reporting. From an evolutionary standpoint, the behaviour correlates of defeats of loss are thought to be an adaptive response to prevent further loss. Therefore, attempts to model depression that seeks to induce defeat or despair may actually reflect adaption and not disease. Furthermore, while depression and anxiety are frequently comorbid, dissociation of the two in animal models is difficult to achieve. Pharmacological assessment of validity is frequently disconnected from clinical pharmacotherapeutics in that most screening tests assess acute effects, while antidepressants normally take a few weeks to work in humans.
Neurocircuits
Regions involved in reward are common targets of manipulation in animal models of depression, including the nucleus accumbens (NAc), ventral tegmental area (VTA), ventral pallidum (VP), lateral habenula (LHb) and medial prefrontal cortex (mPFC). Tentative fMRI studies in humans demonstrate elevated LHb activity in depression. The lateral habenula projects to the RMTg to drive inhibition of dopamine neurons in the VTA during omission of reward. In animal models of depression, elevated activity has been reported in LHb neurons that project to the ventral tegmental area (ostensibly reducing dopamine release). The LHb also projects to aversion reactive mPFC neurons, which may provide an indirect mechanism for producing depressive behaviours. Learned helplessness induced potentiation of LHb synapses are reversed by antidepressant treatment, providing predictive validity. A number of inputs to the LHb have been implicated in producing depressive behaviours. Silencing GABAergic projections from the NAc to the LHb reduces conditioned place preference induced in social aggression, and activation of these terminals induces CPP. Ventral pallidum firing is also elevated by stress induced depression, an effect that is pharmacologically valid, and silencing of these neurons alleviates behavioural correlates of depression. Tentative in vivo evidence from people with MDD suggests abnormalities in dopamine signalling. This led to early studies investigating VTA activity and manipulations in animal models of depression. Massive destruction of VTA neurons enhances depressive behaviours, while VTA neurons reduce firing in response to chronic stress. However, more recent specific manipulations of the VTA produce varying results, with the specific animal model, duration of VTA manipulation, method of VTA manipulation, and subregion of VTA manipulation all potentially leading to differential outcomes. Stress and social defeat induced depressive symptoms, including anhedonia, are associated with potentiation of excitatory inputs to Dopamine D2 receptor-expressing medium spiny neurons (D2-MSNs) and depression of excitatory inputs to Dopamine D1 receptor-expressing medium spiny neurons (D1-MSNs). Optogenetic excitation of D1-MSNs alleviates depressive symptoms and is rewarding, while the same with D2-MSNs enhances depressive symptoms. Excitation of glutaminergic inputs from the ventral hippocampus reduces social interactions, and enhancing these projections produces susceptibility to stress-induced depression. Manipulations of different regions of the mPFC can produce and attenuate depressive behaviours. For example, inhibiting mPFC neurons specifically in the intralimbic cortex attenuates depressive behaviours. The conflicting findings associated with mPFC stimulation, when compared to the relatively specific findings in the infralimbic cortex, suggest that the prelimbic cortex and infralimbic cortex may mediate opposing effects. mPFC projections to the raphe nuclei are largely GABAergic and inhibit the firing of serotonergic neurons. Specific activation of these regions reduce immobility in the forced swim test but do not affect open field or forced swim behaviour. Inhibition of the raphe shifts the behavioural phenotype of uncontrolled stress to a phenotype closer to that of controlled stress.
Altered Neuroplasticity
Recent studies have called attention to the role of altered neuroplasticity in depression. A review found a convergence of three phenomena:
Chronic stress reduces synaptic and dendritic plasticity;
Depressed subjects show evidence of impaired neuroplasticity (e.g. shortening and reduced complexity of dendritic trees); and
Anti-depressant medications may enhance neuroplasticity at both a molecular and dendritic level.
The conclusion is that disrupted neuroplasticity is an underlying feature of depression, and is reversed by antidepressants.
Blood levels of BDNF in people with MDD increase significantly with antidepressant treatment and correlate with decrease in symptoms. Post mortem studies and rat models demonstrate decreased neuronal density in the prefrontal cortex thickness in people with MDD. Rat models demonstrate histological changes consistent with MRI findings in humans, however studies on neurogenesis in humans are limited. Antidepressants appear to reverse the changes in neurogenesis in both animal models and humans.
Inflammation
Various reviews have found that general inflammation may play a role in depression. One meta analysis of cytokines in people with MDD found increased levels of pro-inflammatory IL-6 and TNF-a levels relative to controls. The first theories came about when it was noticed that interferon therapy caused depression in a large number of people receiving it. Meta analysis on cytokine levels in people with MDD have demonstrated increased levels of IL-1, IL-6, C-reactive protein, but not IL-10. Increased numbers of T-Cells presenting activation markers, levels of neopterin, IFN gamma, sTNFR, and IL-2 receptors have been observed in depression. Various sources of inflammation in depressive illness have been hypothesized and include trauma, sleep problems, diet, smoking and obesity. Cytokines, by manipulating neurotransmitters, are involved in the generation of sickness behaviour, which shares some overlap with the symptoms of depression. Neurotransmitters hypothesized to be affected include dopamine and serotonin, which are common targets for antidepressant drugs. Induction of indolamine-2,3 dioxygenease by cytokines has been proposed as a mechanism by which immune dysfunction causes depression. One review found normalization of cytokine levels after successful treatment of depression. A meta analysis published in 2014 found the use of anti-inflammatory drugs such as NSAIDs and investigational cytokine inhibitors reduced depressive symptoms. Exercise can act as a stressor, decreasing the levels of IL-6 and TNF-a and increasing those of IL-10, an anti-inflammatory cytokine.
Inflammation is also intimately linked with metabolic processes in humans. For example, low levels of Vitamin D have been associated with greater risk for depression. The role of metabolic biomarkers in depression is an active research area. Recent work has explored the potential relationship between plasma sterols and depressive symptom severity.
Oxidative Stress
A marker of DNA oxidation, 8-Oxo-2′-deoxyguanosine, has been found to be increased in both the plasma and urine of people with MDD. This along with the finding of increased F2-isoprostanes levels found in blood, urine and cerebrospinal fluid indicate increased damage to lipids and DNA in people with MDD. Studies with 8-Oxo-2′ Deoxyguanosine varied by methods of measurement and type of depression, but F2-Isoprostane level was consistent across depression types. Authors suggested lifestyle factors, dysregulation of the HPA axis, immune system and autonomics nervous system as possible causes. Another meta-analysis found similar results with regards to oxidative damage products as well as decreased oxidative capacity. Oxidative DNA damage may play a role in MDD.
Mitochondrial Dysfunction:
Increased markers of oxidative stress relative to controls have been found in people with MDD. These markers include high levels of RNS and ROS which have been shown to influence chronic inflammation, damaging the electron transport chain and biochemical cascades in mitochondria. This lowers the activity of enzymes in the respiratory chain resulting in mitochondrial dysfunction. The brain is a highly energy-consuming and has little capacity to store glucose as glycogen and so depends greatly on mitochondria. Mitochondrial dysfunction has been linked to the dampened neuroplasticity observed in depressed brains.
Large-Scale Brain Network Theory
Instead of studying one brain region, studying large scale brain networks is another approach to understanding psychiatric and neurological disorders, supported by recent research that has shown that multiple brain regions are involved in these disorders. Understanding the disruptions in these networks may provide important insights into interventions for treating these disorders. Recent work suggests that at least three large-scale brain networks are important in psychopathology.
Central Executive Network
The central executive network is made up of fronto-parietal regions, including dorsolateral prefrontal cortex and lateral posterior parietal cortex. This network is involved in high level cognitive functions such as maintaining and using information in working memory, problem solving, and decision making. Deficiencies in this network are common in most major psychiatric and neurological disorders, including depression. Because this network is crucial for everyday life activities, those who are depressed can show impairment in basic activities like test taking and being decisive.
Default Mode Network
The default mode network includes hubs in the prefrontal cortex and posterior cingulate, with other prominent regions of the network in the medial temporal lobe and angular gyrus. The default mode network is usually active during mind-wandering and thinking about social situations. In contrast, during specific tasks probed in cognitive science (for example, simple attention tasks), the default network is often deactivated. Research has shown that regions in the default mode network (including medial prefrontal cortex and posterior cingulate) show greater activity when depressed participants ruminate (that is, when they engage in repetitive self-focused thinking) than when typical, healthy participants ruminate. People with MDD also show increased connectivity between the default mode network and the subgenual cingulate and the adjoining ventromedial prefrontal cortex in comparison to healthy individuals, individuals with dementia or with autism. Numerous studies suggest that the subgenual cingulate plays an important role in the dysfunction that characterizes major depression. The increased activation in the default mode network during rumination and the atypical connectivity between core default mode regions and the subgenual cingulate may underlie the tendency for depressed individual to get “stuck” in the negative, self-focused thoughts that often characterise depression. However, further research is needed to gain a precise understanding of how these network interactions map to specific symptoms of depression.
Salience Network
The salience network is a cingulate-frontal operculum network that includes core nodes in the anterior cingulate and anterior insula. A salience network is a large-scale brain network involved in detecting and orienting the most pertinent of the external stimuli and internal events being presented. Individuals who have a tendency to experience negative emotional states (scoring high on measures of neuroticism) show an increase in the right anterior insula during decision-making, even if the decision has already been made. This atypically high activity in the right anterior insula is thought to contribute to the experience of negative and worrisome feelings. In MDD, anxiety is often a part of the emotional state that characterises depression.
Clinical neuropsychology is a sub-field of psychology concerned with the applied science of brain-behaviour relationships.
Clinical neuropsychologists use this knowledge in the assessment, diagnosis, treatment, and or rehabilitation of patients across the lifespan with neurological, medical, neurodevelopmental and psychiatric conditions, as well as other cognitive and learning disorders. The branch of neuropsychology associated with children and young people is paediatric neuropsychology.
Clinical neuropsychology is a specialised form of clinical psychology. Strict rules are in place to maintain evidence as a focal point of treatment and research within clinical neuropsychology. The assessment and rehabilitation of neuropsychopathologies is the focus for a clinical neuropsychologist. A clinical neuropsychologist must be able to determine whether a symptom(s) may be caused by an injury to the head through interviewing a patient in order to determine what actions should be taken to best help the patient. Another duty of a clinical neuropsychologist is to find cerebral abnormalities and possible correlations. Evidence based practice in both research and treatment is paramount to appropriate clinical neuropsychological practice.
Assessment is primarily by way of neuropsychological tests, but also includes patient history, qualitative observation and may draw on findings from neuroimaging and other diagnostic medical procedures. Clinical neuropsychology requires an in-depth knowledge of: neuroanatomy, neurobiology, psychopharmacology and neuropathology.
Brief History
During the late 1800s, brain-behaviour relationships were interpreted by European physicians who observed and identified behavioural syndromes that were related with focal brain dysfunction.
Clinical neuropsychology is a fairly new practice in comparison to other specialty fields in psychology with history going back to the 1960s. The specialty focus of clinical neuropsychology evolved slowly into a more defined whole as interest grew. Threads from neurology, clinical psychology, psychiatry, cognitive psychology, and psychometrics all have been woven together to create the intricate tapestry of clinical neuropsychology, a practice which is very much so still evolving. The history of clinical neuropsychology is long and complicated due to its ties to so many older practices. Researchers like Thomas Willis (1621-1675) who has been credited with creating neurology, John Hughlings Jackson (1835-1911) who theorised that cognitive processes occurred in specific parts of the brain, Paul Broca (1824-1880) and Karl Wernicke (1848-1905) who studied the human brain in relation to psychopathology, Jean Martin Charcot (1825-1893) who apprenticed Sigmund Freud (1856-1939) who created the psychoanalytic theory all contributed to clinical medicine which later contributed to clinical neuropsychology. The field of psychometrics contributed to clinical neuropsychology through individuals such as Francis Galton (1822-1911) who collected quantitative data on physical and sensory characteristics, Karl Pearson (1857-1936) who established the statistics which psychology now relies on, Wilhelm Wundt (1832-1920) who created the first psychology lab, his student Charles Spearman (1863-1945) who furthered statistics through discoveries like factor analysis, Alfred Binet (1857-1911) and his apprentice Theodore Simon (1872-1961) who together made the Binet-Simon scale of intellectual development, and Jean Piaget (1896-1980) who studied child development. Studies in intelligence testing made by Lewis Terman (1877-1956) who updated the Binet-Simon scale to the Stanford-Binet intelligence scale, Henry Goddard (1866-1957) who developed different classification scales, and Robert Yerkes (1876-1956) who was in charge of the Army Alpha and Beta tests also all contributed to where clinical neuropsychology is today.
Clinical neuropsychology focuses on the brain and goes back to the beginning of the 20th century. As a clinician a clinical neuropsychologist offers their services by addressing three steps: assessment, diagnosis, and treatment. The term clinical neuropsychologist was first made by Sir William Osler on 16 April 1913. While clinical neuropsychology was not a focus until the 20th century evidence of brain and behaviour treatment and studies are seen as far back as the neolithic area when trephination, a crude surgery in which a piece of the skull is removed, has been observed in skulls. As a profession, clinical neuropsychology is a subspecialty beneath clinical psychology. During World War I (1914-1918) the early term shell shock was first observed in soldiers who survived the war. This was the beginning of efforts to understand traumatic events and how they affected people. During the Great Depression (1929-1941) further stressors caused shell shock like symptoms to emerge. In World War II (1939-1945) the term shell shock was changed to battle fatigue and clinical neuropsychology became even more involved with attempting to solve the puzzle of peoples’ continued signs of trauma and distress. The Veterans Administration or VA was created in 1930 which increased the call for clinical neuropsychologists and by extension the need for training. The Korean War (1950-1953) and Vietnam War (1960-1973) further solidified the need for treatment by trained clinical neuropsychologists. In 1985 the term post-traumatic stress disorder or PTSD was coined and the understanding that traumatic events of all kinds could cause PTSD started to evolve.
The relationship between human behaviour and the brain is the focus of clinical neuropsychology as defined by Meir in 1974. There are two subdivisions of clinical neuropsychology which draw much focus; organic and environmental natures. Ralph M. Reitan, Arthur L. Benton, and A.R. Luria are all past neuropsychologists whom believed and studied the organic nature of clinical neuropsychology. Alexander Luria is the Russian neuropsychologist responsible for the origination of clinical psychoneurological assessment after WWII. Building upon his original contribution connecting the voluntary and involuntary functions influencing behaviour, Luria further conjoins the methodical structures and associations of neurological processes in the brain. Luria developed the ‘combined motor method’ to measure thought processes based on the reaction times when three simultaneous tasks are appointed that require a verbal response. On the other side, environmental nature of clinical neuropsychology did not appear until more recently and is characterised by treatments such as behaviour therapy. The relationship between physical brain abnormalities and the presentation of psychopathology is not completely understood, but this is one of the questions which clinical neuropsychologists hope to answer in time. In 1861 the debate over human potentiality versus localisation began. The two sides argued over how human behaviour presented in the brain. Paul Broca postulated that cognitive problems could be caused by physical damage to specific parts of the brain based on a case study of his in which he found a lesion on the brain of a deceased patient who had presented the symptom of being unable to speak, that portion of the brain is now known as Broca’s Area. In 1874 Carl Wernicke also made a similar observation in a case study involving a patient with a brain lesion whom was unable to comprehend speech, the part of the brain with the lesion is now deemed Wernicke’s Area. Both Broca and Wernicke believed and studied the theory of localisation. On the other hand, equal potentiality theorists believed that brain function was not based on a single piece of the brain but rather on the brain as a whole. Marie J.P Flourens conducted animal studies in which he found that the amount of brain tissue damaged directly affected the amount that behaviour ability was altered or damaged. Kurt Goldstein observed the same idea as Flourens except in veterans who had fought in World War I. In the end, despite all of the disagreement, neither theory completely explains the human brains complexity. Thomas Hughlings Jackson created a theory which was thought to be a possible solution. Jackson believed that both potentiality and localisation were in part correct and that behaviour was made by multiple parts of the brain working collectively to cause behaviours, and Luria (1966-1973) furthered Jackson’s theory.
The Role
When considering where a clinical neuropsychologist works, hospitals are a common place for practitioners to end up. There are three main variations in which a clinical neuropsychologist may work at a hospital; as an employee, consultant, or independent practitioner. As a clinical neuropsychologist working as an employee of a hospital the individual may receive a salary, benefits, and sign a contract for employment. In the case of an employee of a hospital the hospital is in charge of legal and financial responsibilities. The second option of working as a consultant implies that the clinical neuropsychologist is part of a private practice or is a member of a physicians group. In this scenario, the clinical neuropsychologist may work in the hospital like the employee of the hospital but all financial and legal responsibilities go through the group which the clinical neuropsychologist is a part of. The third option is an independent practitioner whom works alone and may even have their office outside of the hospital or rent a room in the hospital. In the third case, the clinical neuropsychologist is completely on their own and in charge of their own financial and legal responsibilities.
Assessment
Assessments are used in clinical neuropsychology to find brain psychopathologies of the cognitive, behavioural, and emotional variety. Physical evidence is not always readily visible so clinical neuropsychologists must rely on assessments to tell them the extent of the damage. The cognitive strengths and weaknesses of the patient are assessed to help narrow down the possible causes of the brain pathology. A clinical neuropsychologist is expected to help educate the patient on what is happening to them so that the patient can understand how to work with their own cognitive deficits and strengths. An assessment should accomplish many goals such as; gage consequences of impairments to quality of life, compile symptoms and the change in symptoms over time, and assess cognitive strengths and weaknesses. Accumulation of the knowledge earned from the assessment is then dedicated to developing a treatment plan based on the patient’s individual needs. An assessment can also help the clinical neuropsychologist gauge the impact of medications and neurosurgery on a patient. Behavioural neurology and neuropsychology tools can be standardised or psychometric tests and observational data collected on the patient to help build an understanding of the patient and what is happening with them. There are essential prerequisites which must be present in a patient in order for the assessment to be effective; concentration, comprehension, and motivation and effort.
Lezak lists six primary reasons neuropsychological assessments are carried out: diagnosis, patient care and its planning, treatment planning, treatment evaluation, research and forensic neuropsychology. To conduct a comprehensive assessment will typically take several hours and may need to be conducted over more than a single visit. Even the use of a screening battery covering several cognitive domains may take 1.5-2 hours. At the commencement of the assessment it is important to establish a good rapport with the patient and ensure they understand the nature and aims of the assessment.
Neuropsychological assessment can be carried out from two basic perspectives, depending on the purpose of assessment. These methods are normative or individual. Normative assessment, involves the comparison of the patient’s performance against a representative population. This method may be appropriate in investigation of an adult onset brain insult such as traumatic brain injury or stroke. Individual assessment may involve serial assessment, to establish whether declines beyond those which are expected to occur with normal aging, as with dementia or another neurodegenerative condition.
Assessment can be further subdivided into sub-sections:
History Taking
Neuropsychological assessments usually commence with a clinical interview as a means of collecting a history, which is relevant to the interpretation of any later neuropsychological tests. In addition, this interview provides qualitative information about the patient’s ability to act in a socially apt manner, organise and communicate information effectively and provide an indication as to the patient’s mood, insight and motivation. It is only within the context of a patient’s history that an accurate interpretation of their test data and thus a diagnosis can be made. The clinical interview should take place in a quiet area free from distractions. Important elements of a history include demographic information, description of presenting problem, medical history (including any childhood or developmental problems, psychiatric and psychological history), educational and occupational history (and if any legal history and military history).
Selection of Neuropsychological Tests
It is not uncommon for patients to be anxious about being tested; explaining that tests are designed so that they will challenge everyone and that no one is expected to answer all questions correctly may be helpful. An important consideration of any neuropsychological assessment is a basic coverage of all major cognitive functions. The most efficient way to achieve this is the administration of a battery of tests covering: attention, visual perception and reasoning, learning and memory, verbal function, construction, concept formation, executive function, motor abilities and emotional status. Beyond this basic battery, choices of neuropsychological tests to be administered are mainly made on the basis of which cognitive functions need to be evaluated in order to fulfil the assessment objectives.
Report Writing
Following a neuropsychological assessment it is important to complete a comprehensive report based on the assessment conducted. The report is for other clinicians, as well as the patient and their family so it is important to avoid jargon or the use of language which has different clinical and lay meanings (e.g. intellectually disabled as the correct clinical term for an IQ below 70, but offensive in lay language). The report should cover background to the referral, relevant history, reasons for assessment, neuropsychologists observations of patient’s behaviour, test administered and results for cognitive domains tested, any additional findings (e.g. questionnaires for mood) and finish the report with a summary and recommendations. In the summary it is important to comment on what the profile of results indicates regarding the referral question. The recommendations section contains practical information to assist the patient and family, or improve the management of the patient’s condition.
Educational Requirements of Different Countries
The educational requirements for becoming a clinical neuropsychologist differ between countries. In some countries it may be necessary to complete a clinical psychology degree, before specialising with further studies in clinical neuropsychology. While some countries offer clinical neuropsychology courses to students who have completed 4 years of psychology studies. All clinical neuropsychologists require a postgraduate qualification, whether it be a Masters or Doctorate (Ph.D, Psy.D. or D.Psych).
Australia
To become a clinical neuropsychologist in Australia requires the completion of a 3-year Australian Psychology Accreditation Council (APAC) approved undergraduate degree in psychology, a 1-year psychology honours, followed by a 2-year Masters or 3-year Doctorate of Psychology (D.Psych) in clinical neuropsychology. These courses involve coursework (lectures, tutorials, practicals etc.), supervised practice placements and the completion of a research thesis. Masters and D.Psych courses involve the same amount of coursework units, but differ in the amount of supervised placements undertaken and length of research thesis. Masters courses require a minimum of 1,000 hours (125 days) and D.Psych courses require a minimum of 1,500 hours (200 days), it is mandatory that these placements expose students to acute neurology/neurosurgery, rehabilitation, psychiatric, geriatric and paediatric populations.
Canada
To become a clinical neuropsychologist in Canada requires the completion of a 4-year honours degree in psychology and a 4-year doctoral degree in clinical neuropsychology. Often a 2-year master’s degree is required before commencing the doctoral degree. The doctoral degree involves coursework and practical experience (practicum and internship). Practicum is between 600 and 1,000 hours of practical application of skills acquired in the programme. At least 300 hours must be supervised, face-to-face client contact. The practicum is intended to prepare students for the internship/residency. Internships/residencies are a year long experience in which the student functions as a neuropsychologist, under supervision. Currently, there are 3 CPA-accredited Clinical Neuropsychology internships/residencies in Canada, although other unaccredited ones exist. Prior to commencing the internship students must have completed all doctoral coursework, received approval for their thesis proposal (if not completed the thesis) and the 600 hours of practicum.
United Kingdom
To become a clinical neuropsychologist in the UK, requires prior qualification as a clinical or educational psychologist as recognised by the Health Professions Council, followed by further postgraduate study in clinical neuropsychology. In its entirety, education to become a clinical neuropsychologist in the UK consists of the completion of a 3-year British Psychological Society accredited undergraduate degree in psychology, 3-year Doctorate in clinical (usually D.Clin.Psy.) or educational psychology (D.Ed.Psy.), followed by a 1-year Masters (MSc) or 9-month Postgraduate Diploma (PgDip) in Clinical Neuropsychology. The British Psychological Division of Counselling Psychology are also currently offering training to its members in order to ensure that they can apply to be registered Neuropsychologists also.
United States
In order to become a clinical neuropsychologist in the US and be compliant with Houston Conference Guidelines, the completion of a 4-year undergraduate degree in psychology and a 4 to 5-year doctoral degree (Psy.D. or Ph.D.) must be completed. After the completion of the doctoral coursework, training and dissertation, students must complete a 1-year internship, followed by an additional 2 years of supervised residency. The doctoral degree, internship and residency must all be undertaken at American Psychological Association approved institutions. After the completion of all training, students must apply to become licensed in their state to practice psychology. The American Board of Clinical Neuropsychology, The American Board of Professional Neuropsychology, and The American Board of Paediatric Neuropsychology all award board certification to neuropsychologists that demonstrate competency in specific areas of neuropsychology, by reviewing the neuropsychologist’s training, experience, submitted case samples, and successfully completing both written and oral examinations. Although these requirements are standard according to Houston Conference Guidelines, even these guidelines have stated that the completion of all of these requirements is still aspirational, and other ways of achieving clinical neuropsychologist status are possible.
Neuropsychoanalysis integrates both neuroscience and psychoanalysis, to create a balanced and equal study of the human mind.
This overarching approach began as advances in neuroscience lead to breakthroughs which held pertinent information for the field of psychoanalysis. Despite advantages for these fields to interconnect, there is some concern that too much emphasis on neurobiological physiology of the brain will undermine the importance of dialogue and exploration that is foundational to the field of psychoanalysis. Critics will also point to the qualitative and subjective nature of the field of psychoanalysis, claiming it cannot be fully reconciled with the quantitative and objective nature of neuroscientific research.
However, despite this critique, proponents of the field of neuropsychoanalysis remind critics that the father of psychoanalysis, Sigmund Freud himself, began his career as a neuroanatomist, further arguing that research in this category proves that the psychodynamic effects of the mind are inextricably linked to neural activity in the brain. Indeed, neuroscientific progress has created a shared study of many of the same cognitive phenomenon, and proponents for a distinct field under the heading of neuropsychoanalysis point to the ability for observation of both the subjective mind and empirical evidence in neurobiology to provide greater understanding and greater curative methods.
Therefore, neurospsychoanalysis aims to bring a field, often viewed as belonging more to the humanities than the sciences, into the scientific realm and under the umbrella of neuroscience, distinct from psychoanalysis, and yet adding to the plethora of insight garnered from it.
Brief History
Neuropsychoanalysis as a discipline can be traced as far back as Sigmund Freud’s manuscript, “Project for a Scientific Psychology”. Written in 1895, but only published posthumously, Freud developed his theories of the neurobiological function of the storage of memory in this work. His statement, based on his theory that memory is biologically stored in the brain by, “a permanent alteration following an event”, had a prophetic insight into the empirical discoveries that would corroborate these theories close to 100 years later. Freud speculated that psychodynamics and neurobiology would eventually reunite as one field of study. While time would eventually prove him correct to some degree, the latter half of the 20th century only saw a very gradual movement in this direction with only a few individuals championing this line of thought.
Significant advances in neuroscience throughout the 20th century created a clearer understanding of the functionality of the brain, which have vastly enhanced the way we view the mind. This began in the 1930s with the invention of electroencephalography, which enabled imaging of the brain as never seen before. A decade later the use of dynamic localisation, or the lesion method, further shed light onto the interaction of systems in the brain. Computerised tomography (CT) lead to even greater understanding of the interaction within the brain, and finally the invention of multiple scan technologies in the 1990s, the functional magnetic resonance imaging (fMRI), positron emission tomography (PET), and the single photon emission computed tomography (SPECT) gave researchers empirical evidence of neurobiological processes.
It was in 1999, just before the turn of the century, that the term “neuropsychoanalysis” was used in a new journal entitled with the same name. This term once was hyphenated to indicate that the conjoining of the two fields of study did not suggest that they had been fully integrated, but rather that this new line of scientific inquiry was interdisciplinary. With repeated use, the hyphen was lost, and the name appears as we see it today.
Theoretical Base
Dual-Aspect Monism
Neuropsychoanalysis is best described as a marriage between neuroscience and psychoanalysis. However, its relationship to the broader field of neuropsychology – which relates the biological brain to psychological functions and behaviour – cannot be denied. Indeed, neuropsychoanalysis further seeks to remedy classical neurology’s exclusion of the subjective mind.
The subjective mind, that is, sensations, thoughts, feelings and consciousness, can seem antithetical to the cellular matter that makes up the neurobiology of the brain. Indeed, while Freud is most often credited with being the seminal creator of the study of the mind in modern terms, it was Descartes who concluded that mind and brain were two entirely different kinds of stuff. Accordingly, he invented the “dualism” of the mind, the mind-body dichotomy. Body is one kind of thing, and mind (or spirit or soul) is another. But since this second kind of stuff does not lend itself to scientific inquiry, many of today’s psychologists and neuroscientists have seemingly rejected Cartesian dualism.
Neuropsychoanalysis meets this challenge via dual-aspect monism, sometimes referred to as perspectivism. That is, we are monistic. Our brains, including mind, are made of one kind of stuff, cells, but we perceive this stuff in two different ways.
Psychoanalysis as a Foundation
Perhaps because Freud himself began his career as a neurologist, psychoanalysis has given the field of neuroscience the platform upon which many of its scientific hypotheses were founded. With the field of psychoanalysis suffering from what many see as a decline in innovation and popularity, a call for new approaches and a more scientific methodology is long overdue. The history of neuropsychoanalysis therefore, goes some way in explaining why some consider it the logical conclusion, and representative of an evolution that psychoanalysis was in need of. Since the mind itself is viewed as purely ontological, our appreciation of reality is dependent on neurobiological functions of the brain, which we can use to observe “subjectively,” from inside, how we feel and what we think. Freud refined this kind of observation into free association. He claimed and that this is the best technique that we have for perceiving complex mental functions that simple introspection will not reveal. Through psychoanalysis, we can discover mind’s unconscious functioning.
Neuroscience as a Foundation
Due to the very nature of neuropsychoanalysis, those working in this burgeoning field have been able to draw useful insights from a number of distinguished neuroscientists, indeed many of these now serve on the editorial board of the journal Neuropsychoanalysis. Some of these more notable names foundational to the development of neuropsychoanalysis include:
Antonio Damasio.
Eric Kandel.
Joseph LeDoux.
Helen Mayberg.
Jaak Panksepp.
VS Ramachandran.
Oliver Sacks.
Mark Solms.
Neuroscientists, often studying the same cognitive functions of the brain as psychoanalysts, do so in quantitative methods such as dissection post mortem, small lesions administered to create certain curative effects, or with the visual and objective aid of brain imaging, all of which enable researchers to trace neurochemical pathways and build a more accurate understanding of the physical functioning of the brain. Another branch of neuroscience also observes the “mind” from outside, that is, by means of neurological examination. This is often done in the form of physical tests, such as questionnaires, the Boston Naming test or Wisconsin Sorting, creating bisecting lines, acting out how one performs daily tasks such as a screwdriver, just to name a few. Neurologists can compare the changes in psychological function that the neurological examination shows with the associated changes in the brain, either post mortem or by means of modern imaging technology. Much of neuroscience aims to break down and tease out the cognitive and biological functions behind both conscious and unconscious actions within the brain. In this way it is no different than psychoanalysis, which has had similar goals since its inception. Therefore, to ignore the additional insight neuroscience can offer psychoanalysis would be to limit a huge source of knowledge that can only enhance psychoanalysis as a whole.
Models of Pathologies
Depression
Heinz Böker and Rainer Krähenman proposed a model depression as dysregulation of the relationship between the self and the other. This psychodynamic model, is related to the neurobiological model of the default mode network, DMN, and the executive network, EN, of the brain, noting experimentally the DMN seemed to be more active in depressed patients. The psychological construct of rumination is conceptualised which is experimentally more common in depressed patients, is viewed as equivalent to the cognitive processing of the self, and therefore the activation of the DMN. Similarly, experimentally measurable constructs of attribution bias are viewed as being related to this “cognitive processing of self”. It has been shown that forms of psychodynamic therapy for depression have effects on the activation of several areas of the brain.
Research Directions
Neuropsychoanalytic relate unconscious (and sometimes conscious) functioning discovered through the techniques of psychoanalysis or experimental psychology to underlying brain processes. Among the ideas explored in recent research are the following:
“Consciousness” is limited (5-9 bits of information) compared to emotional and unconscious thinking based in the limbic system.
Note: Solm’s book showed as reference in the footnote does not provide such an information.
It may be confused with the capacity of short-term memory.
Secondary-process, reality-oriented thinking can be understood as frontal lobe executive control systems.
Dreams, confabulations, and other expressions of primary-process thinking are meaningful, wish-fulfilling manifestations of the loss of frontal executive control of mesocortical and mesolimbic “seeking” systems.
Freud’s “libido” corresponds to a dopaminergic seeking system.
Drives can be understood as a series of basic emotions (prompts to action) anchored in pontine regions, specifically the periaqueductal gray, and projecting to cortex: play; seeking; caring; fear; anger; sadness. Seeking is constantly active; the others seek appropriate consummations (corresponding to Freud’s “dynamic” unconscious).
Seemingly rational and conscious decisions are driven from the limbic system by emotions which are unconscious.
Infantile amnesia (the absence of memory for the first years of life) occurs because the verbal left hemisphere becomes activated later, in the second or third year of life, after the non-verbal right hemisphere.
But infants can and do have procedural and emotional memories.
Infants’ first-year experiences of attachment and second-year (approximately) experiences of disapproval lay down pathways that regulate emotions and profoundly affect adult personality.
Oedipal behaviors (observable in primates) can be understood as the effort to integrate lust systems (testosterone-driven), romantic love (dopamine-driven), and attachment (oxytocin-driven) in relation to key persons in the environment.
Differences between the sexes are more biologically-based and less environmentally-driven than Freud believed.
Neurobiologically Informed Trauma Therapy with Children and Adolescents: Understanding Mechanisms of Change (Norton Series on Interpersonal Neurobiology).
Author(s): Linda Chapman.
Year: 2014.
Edition: First (1st).
Publisher: W.W. Norton & Company.
Type(s): Paperback and Kindle.
Synopsis:
The model of treatment developed here is grounded in the physical, psychological, and cognitive reactions children have to traumatic experiences and the consequences of those experiences. The approach to treatment utilises the integrative capacity of the brain to create a self, foster insight, and produce change. Treatment strategies are based on cutting-edge understanding of neurobiology, the development of the brain, and the storage and retrieval of traumatic memory. Case vignettes illustrate specific examples of the reactions of children, families, and teens to acute and repeated exposure to traumatic events.
Also presented is the most recent knowledge of the role of the right hemisphere (RH) in development and therapy. Right brain communication, and how to recognise the non-verbal symbolic and unconscious, affective processes will be explained, along with examples of how the therapist can utilise art making, media, tools, and self to engage in a two-person biology. 30 illustrations; 8 pages of colour.
Neuroscience (or neurobiology) is the scientific study of the nervous system. It is a multidisciplinary science that combines physiology, anatomy, molecular biology, developmental biology, cytology, mathematical modelling, and psychology to understand the fundamental and emergent properties of neurons and neural circuits. The understanding of the biological basis of learning, memory, behaviour, perception, and consciousness has been described by Eric Kandel as the “ultimate challenge” of the biological sciences.
The scope of neuroscience has broadened over time to include different approaches used to study the nervous system at different scales and the techniques used by neuroscientists have expanded enormously, from molecular and cellular studies of individual neurons to imaging of sensory, motor and cognitive tasks in the brain.
Brief History
The earliest study of the nervous system dates to ancient Egypt. Trepanation, the surgical practice of either drilling or scraping a hole into the skull for the purpose of curing head injuries or mental disorders, or relieving cranial pressure, was first recorded during the Neolithic period. Manuscripts dating to 1700 BC indicate that the Egyptians had some knowledge about symptoms of brain damage.
Early views on the function of the brain regarded it to be a “cranial stuffing” of sorts. In Egypt, from the late Middle Kingdom onwards, the brain was regularly removed in preparation for mummification. It was believed at the time that the heart was the seat of intelligence. According to Herodotus, the first step of mummification was to “take a crooked piece of iron, and with it draw out the brain through the nostrils, thus getting rid of a portion, while the skull is cleared of the rest by rinsing with drugs.”
The view that the heart was the source of consciousness was not challenged until the time of the Greek physician Hippocrates. He believed that the brain was not only involved with sensation – since most specialised organs (e.g. eyes, ears, tongue) are located in the head near the brain – but was also the seat of intelligence. Plato also speculated that the brain was the seat of the rational part of the soul. Aristotle, however, believed the heart was the centre of intelligence and that the brain regulated the amount of heat from the heart. This view was generally accepted until the Roman physician Galen, a follower of Hippocrates and physician to Roman gladiators, observed that his patients lost their mental faculties when they had sustained damage to their brains.
Abulcasis, Averroes, Avicenna, Avenzoar, and Maimonides, active in the Medieval Muslim world, described a number of medical problems related to the brain. In Renaissance Europe, Vesalius (1514-1564), René Descartes (1596-1650), Thomas Willis (1621-1675) and Jan Swammerdam (1637-1680) also made several contributions to neuroscience.
Luigi Galvani’s pioneering work in the late 1700s set the stage for studying the electrical excitability of muscles and neurons. In the first half of the 19th century, Jean Pierre Flourens pioneered the experimental method of carrying out localised lesions of the brain in living animals describing their effects on motricity, sensibility and behaviour. In 1843 Emil du Bois-Reymond demonstrated the electrical nature of the nerve signal, whose speed Hermann von Helmholtz proceeded to measure, and in 1875 Richard Caton found electrical phenomena in the cerebral hemispheres of rabbits and monkeys. Adolf Beck published in 1890 similar observations of spontaneous electrical activity of the brain of rabbits and dogs. Studies of the brain became more sophisticated after the invention of the microscope and the development of a staining procedure by Camillo Golgi during the late 1890s. The procedure used a silver chromate salt to reveal the intricate structures of individual neurons. His technique was used by Santiago Ramón y Cajal and led to the formation of the neuron doctrine, the hypothesis that the functional unit of the brain is the neuron. Golgi and Ramón y Cajal shared the Nobel Prize in Physiology or Medicine in 1906 for their extensive observations, descriptions, and categorizations of neurons throughout the brain.
In parallel with this research, work with brain-damaged patients by Paul Broca suggested that certain regions of the brain were responsible for certain functions. At the time, Broca’s findings were seen as a confirmation of Franz Joseph Gall’s theory that language was localised and that certain psychological functions were localised in specific areas of the cerebral cortex. The localisation of function hypothesis was supported by observations of epileptic patients conducted by John Hughlings Jackson, who correctly inferred the organisation of the motor cortex by watching the progression of seizures through the body. Carl Wernicke further developed the theory of the specialisation of specific brain structures in language comprehension and production. Modern research through neuroimaging techniques, still uses the Brodmann cerebral cytoarchitectonic map (referring to study of cell structure) anatomical definitions from this era in continuing to show that distinct areas of the cortex are activated in the execution of specific tasks.
During the 20th century, neuroscience began to be recognised as a distinct academic discipline in its own right, rather than as studies of the nervous system within other disciplines. Eric Kandel and collaborators have cited David Rioch, Francis O. Schmitt, and Stephen Kuffler as having played critical roles in establishing the field. Rioch originated the integration of basic anatomical and physiological research with clinical psychiatry at the Walter Reed Army Institute of Research, starting in the 1950s. During the same period, Schmitt established a neuroscience research programme within the Biology Department at the Massachusetts Institute of Technology, bringing together biology, chemistry, physics, and mathematics. The first freestanding neuroscience department (then called Psychobiology) was founded in 1964 at the University of California, Irvine by James L. McGaugh. This was followed by the Department of Neurobiology at Harvard Medical School, which was founded in 1966 by Stephen Kuffler.
The understanding of neurons and of nervous system function became increasingly precise and molecular during the 20th century. For example, in 1952, Alan Lloyd Hodgkin and Andrew Huxley presented a mathematical model for transmission of electrical signals in neurons of the giant axon of a squid, which they called “action potentials”, and how they are initiated and propagated, known as the Hodgkin-Huxley model. In 1961–1962, Richard FitzHugh and J. Nagumo simplified Hodgkin-Huxley, in what is called the FitzHugh-Nagumo model. In 1962, Bernard Katz modelled neurotransmission across the space between neurons known as synapses. Beginning in 1966, Eric Kandel and collaborators examined biochemical changes in neurons associated with learning and memory storage in Aplysia. In 1981 Catherine Morris and Harold Lecar combined these models in the Morris-Lecar model. Such increasingly quantitative work gave rise to numerous biological neuron models and models of neural computation.
As a result of the increasing interest about the nervous system, several prominent neuroscience organizations have been formed to provide a forum to all neuroscientist during the 20th century. For example, the International Brain Research Organisation was founded in 1961, the International Society for Neurochemistry in 1963, the European Brain and Behaviour Society in 1968, and the Society for Neuroscience in 1969. Recently, the application of neuroscience research results has also given rise to applied disciplines as neuroeconomics, neuroeducation, neuroethics, and neurolaw.
Over time, brain research has gone through philosophical, experimental, and theoretical phases, with work on brain simulation predicted to be important in the future.
Modern Neuroscience
The scientific study of the nervous system increased significantly during the second half of the twentieth century, principally due to advances in molecular biology, electrophysiology, and computational neuroscience. This has allowed neuroscientists to study the nervous system in all its aspects: how it is structured, how it works, how it develops, how it malfunctions, and how it can be changed.
For example, it has become possible to understand, in much detail, the complex processes occurring within a single neuron. Neurons are cells specialised for communication. They are able to communicate with neurons and other cell types through specialised junctions called synapses, at which electrical or electrochemical signals can be transmitted from one cell to another. Many neurons extrude a long thin filament of axoplasm called an axon, which may extend to distant parts of the body and are capable of rapidly carrying electrical signals, influencing the activity of other neurons, muscles, or glands at their termination points. A nervous system emerges from the assemblage of neurons that are connected to each other.
The vertebrate nervous system can be split into two parts: the central nervous system (defined as the brain and spinal cord), and the peripheral nervous system. In many species – including all vertebrates – the nervous system is the most complex organ system in the body, with most of the complexity residing in the brain. The human brain alone contains around one hundred billion neurons and one hundred trillion synapses; it consists of thousands of distinguishable substructures, connected to each other in synaptic networks whose intricacies have only begun to be unravelled. At least one out of three of the approximately 20,000 genes belonging to the human genome is expressed mainly in the brain.
Due to the high degree of plasticity of the human brain, the structure of its synapses and their resulting functions change throughout life.
Making sense of the nervous system’s dynamic complexity is a formidable research challenge. Ultimately, neuroscientists would like to understand every aspect of the nervous system, including how it works, how it develops, how it malfunctions, and how it can be altered or repaired. Analysis of the nervous system is therefore performed at multiple levels, ranging from the molecular and cellular levels to the systems and cognitive levels. The specific topics that form the main foci of research change over time, driven by an ever-expanding base of knowledge and the availability of increasingly sophisticated technical methods. Improvements in technology have been the primary drivers of progress. Developments in electron microscopy, computer science, electronics, functional neuroimaging, and genetics and genomics have all been major drivers of progress.
Molecular and Cellular Neuroscience
Basic questions addressed in molecular neuroscience include the mechanisms by which neurons express and respond to molecular signals and how axons form complex connectivity patterns. At this level, tools from molecular biology and genetics are used to understand how neurons develop and how genetic changes affect biological functions. The morphology, molecular identity, and physiological characteristics of neurons and how they relate to different types of behaviour are also of considerable interest.
Questions addressed in cellular neuroscience include the mechanisms of how neurons process signals physiologically and electrochemically. These questions include how signals are processed by neurites and somas and how neurotransmitters and electrical signals are used to process information in a neuron. Neurites are thin extensions from a neuronal cell body, consisting of dendrites (specialised to receive synaptic inputs from other neurons) and axons (specialised to conduct nerve impulses called action potentials). Somas are the cell bodies of the neurons and contain the nucleus.
Another major area of cellular neuroscience is the investigation of the development of the nervous system. Questions include the patterning and regionalisation of the nervous system, neural stem cells, differentiation of neurons and glia (neurogenesis and gliogenesis), neuronal migration, axonal and dendritic development, trophic interactions, and synapse formation.
Computational neurogenetic modelling is concerned with the development of dynamic neuronal models for modelling brain functions with respect to genes and dynamic interactions between genes.
Neural Circuits and Systems
Questions in systems neuroscience include how neural circuits are formed and used anatomically and physiologically to produce functions such as reflexes, multisensory integration, motor coordination, circadian rhythms, emotional responses, learning, and memory. In other words, they address how these neural circuits function in large-scale brain networks, and the mechanisms through which behaviours are generated. For example, systems level analysis addresses questions concerning specific sensory and motor modalities: how does vision work? How do songbirds learn new songs and bats localize with ultrasound? How does the somatosensory system process tactile information? The related fields of neuroethology and neuropsychology address the question of how neural substrates underlie specific animal and human behaviours. Neuroendocrinology and psychoneuroimmunology examine interactions between the nervous system and the endocrine and immune systems, respectively. Despite many advancements, the way that networks of neurons perform complex cognitive processes and behaviours is still poorly understood.
Cognitive and Behavioural Neuroscience
Cognitive neuroscience addresses the questions of how psychological functions are produced by neural circuitry. The emergence of powerful new measurement techniques such as neuroimaging (e.g. fMRI, PET, SPECT), EEG, MEG, electrophysiology, optogenetics and human genetic analysis combined with sophisticated experimental techniques from cognitive psychology allows neuroscientists and psychologists to address abstract questions such as how cognition and emotion are mapped to specific neural substrates. Although many studies still hold a reductionist stance looking for the neurobiological basis of cognitive phenomena, recent research shows that there is an interesting interplay between neuroscientific findings and conceptual research, soliciting and integrating both perspectives. For example, neuroscience research on empathy solicited an interesting interdisciplinary debate involving philosophy, psychology and psychopathology. Moreover, the neuroscientific identification of multiple memory systems related to different brain areas has challenged the idea of memory as a literal reproduction of the past, supporting a view of memory as a generative, constructive and dynamic process.
Neuroscience is also allied with the social and behavioural sciences as well as nascent interdisciplinary fields such as neuroeconomics, decision theory, social neuroscience, and neuromarketing to address complex questions about interactions of the brain with its environment. A study into consumer responses for example uses EEG to investigate neural correlates associated with narrative transportation into stories about energy efficiency.
Computational Neuroscience
Questions in computational neuroscience can span a wide range of levels of traditional analysis, such as development, structure, and cognitive functions of the brain. Research in this field utilises mathematical models, theoretical analysis, and computer simulation to describe and verify biologically plausible neurons and nervous systems. For example, biological neuron models are mathematical descriptions of spiking neurons which can be used to describe both the behaviour of single neurons as well as the dynamics of neural networks. Computational neuroscience is often referred to as theoretical neuroscience.
Nanoparticles in medicine are versatile in treating neurological disorders showing promising results in mediating drug transport across the blood brain barrier. Implementing nanoparticles in antiepileptic drugs enhances their medical efficacy by increasing bioavailability in the bloodstream, as well as offering a measure of control in release time concentration. Although nanoparticles can assist therapeutic drugs by adjusting physical properties to achieve desirable effects, inadvertent increases in toxicity often occur in preliminary drug trials. Furthermore, production of nanomedicine for drug trials is economically consuming, hindering progress in their implementation. Computational models in nanoneuroscience provide alternatives to study the efficacy of nanotechnology-based medicines in neurological disorders while mitigating potential side effects and development costs.
Nanomaterials often operate at length scales between classical and quantum regimes. Due to the associated uncertainties at the length scales that nanomaterials operate, it is difficult to predict their behaviour prior to in vivo studies. Classically, the physical processes which occur throughout neurons are analogous to electrical circuits. Designers focus on such analogies and model brain activity as a neural circuit. Success in computational modelling of neurons have led to the development of stereochemical models that accurately predict acetylcholine receptor-based synapses operating at microsecond time scales.
Ultrafine nanoneedles for cellular manipulations are thinner than the smallest single walled carbon nanotubes. Computational quantum chemistry is used to design ultrafine nanomaterials with highly symmetrical structures to optimise geometry, reactivity and stability.
Behaviour of nanomaterials are dominated by long ranged non-bonding interactions. Electrochemical processes that occur throughout the brain generate an electric field which can inadvertently affect the behaviour of some nanomaterials. Molecular dynamics simulations can mitigate the development phase of nanomaterials as well as prevent neural toxicity of nanomaterials following in vivo clinical trials. Testing nanomaterials using molecular dynamics optimizes nano characteristics for therapeutic purposes by testing different environment conditions, nanomaterial shape fabrications, nanomaterial surface properties, etc without the need for in vivo experimentation. Flexibility in molecular dynamic simulations allows medical practitioners to personalise treatment. Nanoparticle related data from translational nanoinformatics links neurological patient specific data to predict treatment response.
Nano-Neurotechnology
The visualization of neuronal activity is of key importance in the study of neurology. Nano-imaging tools with nanoscale resolution help in these areas. These optical imaging tools are PALM and STORM which helps visualise nanoscale objects within cells. Pampaloni states that, so far, these imaging tools revealed the dynamic behaviour and organisation of the actin cytoskeleton inside the cells, which will assist in understanding how neurons probe their involvement during neuronal outgrowth and in response to injury, and how they differentiate axonal processes and characterisation of receptor clustering and stoichiometry at the plasma inside the synapses, which are critical for understanding how synapses respond to changes in neuronal activity. These past works focused on devices for stimulation or inhibition of neural activity, but the crucial aspect is the ability for the device to simultaneously monitor neural activity. The major aspect that is to be improved in the nano imaging tools is the effective collection of the light as a major problem is that biological tissue are dispersive media that do not allow a straightforward propagation and control of light. These devices use nanoneedle and nanowire (NWs) for probing and stimulation.
NWs are artificial nano- or micro-sized “needles” that can provide high-fidelity electrophysiological recordings if used as microscopic electrodes for neuronal recordings. NWs are an attractive as they are highly functional structures that offer unique electronic properties that are affected by biological/chemical species adsorbed on their surface; mostly the conductivity. This conductivity variance depending on chemical species present allows enhanced sensing performances. NWs are also able to act as non-invasive and highly local probes. These versatility of NWs makes it optimal for interfacing with neurons due to the fact that the contact length along the axon (or the dendrite projection crossing a NW) is just about 20 nm.
Neuroscience and Medicine
Neurology, psychiatry, neurosurgery, psychosurgery, anesthesiology and pain medicine, neuropathology, neuroradiology, ophthalmology, otolaryngology, clinical neurophysiology, addiction medicine, and sleep medicine are some medical specialties that specifically address the diseases of the nervous system. These terms also refer to clinical disciplines involving diagnosis and treatment of these diseases.
Neurology works with diseases of the central and peripheral nervous systems, such as amyotrophic lateral sclerosis (ALS) and stroke, and their medical treatment. Psychiatry focuses on affective, behavioural, cognitive, and perceptual disorders. Anaesthesiology focuses on perception of pain, and pharmacologic alteration of consciousness. Neuropathology focuses upon the classification and underlying pathogenic mechanisms of central and peripheral nervous system and muscle diseases, with an emphasis on morphologic, microscopic, and chemically observable alterations. Neurosurgery and psychosurgery work primarily with surgical treatment of diseases of the central and peripheral nervous systems.
Translational Research
Recently, the boundaries between various specialties have blurred, as they are all influenced by basic research in neuroscience. For example, brain imaging enables objective biological insight into mental illnesses, which can lead to faster diagnosis, more accurate prognosis, and improved monitoring of patient progress over time.
Integrative neuroscience describes the effort to combine models and information from multiple levels of research to develop a coherent model of the nervous system. For example, brain imaging coupled with physiological numerical models and theories of fundamental mechanisms may shed light on psychiatric disorders.
Nanoneuroscience
One of the main goals of nanoneuroscience is to gain a detailed understanding of how the nervous system operates and, thus, how neurons organise themselves in the brain. Consequently, creating drugs and devices that are able to cross the blood brain barrier (BBB) are essential to allow for detailed imaging and diagnoses. The blood brain barrier functions as a highly specialised semipermeable membrane surrounding the brain, preventing harmful molecules that may be dissolved in the circulation blood from entering the central nervous system.
The main two hurdles for drug-delivering molecules to access the brain are size (must have a molecular weight < 400 Da) and lipid solubility. Physicians hope to circumvent difficulties in accessing the central nervous system through viral gene therapy. This often involves direct injection into the patient’s brain or cerebral spinal fluid. The drawback of this therapy is that it is invasive and carries a high risk factor due to the necessity of surgery for the treatment to be administered. Because of this, only 3.6% of clinical trials in this field have progressed to stage III since the concept of gene therapy was developed in the 1980s.
Another proposed way to cross the BBB is through temporary intentional disruption of the barrier. This method was first inspired by certain pathological conditions that were discovered to break down this barrier by themselves, such as Alzheimer’s disease, Parkinson’s disease, stroke, and seizure conditions.
Nanoparticles are unique from macromolecules because their surface properties are dependent on their size, allowing for strategic manipulation of these properties (or, “programming”) by scientists that would not be possible otherwise. Likewise, nanoparticle shape can also be varied to give a different set of characteristics based on the surface area to volume ratio of the particle.
Nanoparticles have promising therapeutic effects when treating neurodegenerative diseases. Oxygen reactive polymer (ORP) is a nano-platform programmed to react with oxygen and has been shown to detect and reduce the presence of reactive oxygen species (ROS) formed immediately after traumatic brain injuries. Nanoparticles have also been employed as a “neuroprotective” measure, as is the case with Alzheimer’s disease and stroke models. Alzheimer’s disease results in toxic aggregates of the amyloid beta protein formed in the brain. In one study, gold nanoparticles were programmed to attach themselves to these aggregates and were successful in breaking them up. Likewise, with ischemic stroke models, cells in the affected region of the brain undergo apoptosis, dramatically reducing blood flow to important parts of the brain and often resulting in death or severe mental and physical changes. Platinum nanoparticles have been shown to act as ROS, serving as “biological antioxidants” and significantly reducing oxidation in the brain as a result of stroke. Nanoparticles can also lead to neurotoxicity and cause permanent BBB damage either from brain oedema or from unrelated molecules crossing the BBB and causing brain damage. This proves further long term in vivo studies are needed to gain enough understanding to allow for successful clinical trials.
One of the most common nano-based drug delivery platforms is liposome-based delivery. They are both lipid-soluble and nano-scale and thus are permitted through a fully functioning BBB. Additionally, lipids themselves are biological molecules, making them highly biocompatible, which in turn lowers the risk of cell toxicity. The bilayer that is formed allows the molecule to fully encapsulate any drug, protecting it while it is travelling through the body. One drawback to shielding the drug from the outside cells is that it no longer has specificity, and requires coupling to extra antibodies to be able to target a biological site. Due to their low stability, liposome-based nanoparticles for drug delivery have a short shelf life.
Targeted therapy using magnetic nanoparticles (MNPs) is also a popular topic of research and has led to several stage III clinical trials. Invasiveness is not an issue here because a magnetic force can be applied from the outside of a patient’s body to interact and direct the MNPs. This strategy has been proven successful in delivering Brain-derived neurotropic factor, a naturally occurring gene thought to promote neurorehabilitation, across the BBB.
Major Branches
Modern neuroscience education and research activities can be very roughly categorised into the following major branches, based on the subject and scale of the system in examination as well as distinct experimental or curricular approaches. Individual neuroscientists, however, often work on questions that span several distinct subfields.
Branch
Description
Affective Neuroscience
Affective neuroscience is the study of the neural mechanisms involved in emotion, typically through experimentation on animal models.
Behavioural Neuroscience
Behavioural neuroscience (also known as biological psychology, physiological psychology, biopsychology, or psychobiology) is the application of the principles of biology to the study of genetic, physiological, and developmental mechanisms of behaviour in humans and non-human animals.
Cellular Neuroscience
Cellular neuroscience is the study of neurons at a cellular level including morphology and physiological properties.
Clinical Neuroscience
The scientific study of the biological mechanisms that underlie the disorders and diseases of the nervous system.
Cognitive Neuroscience
Cognitive neuroscience is the study of the biological mechanisms underlying cognition.
Computational Neuroscience
Computational neuroscience is the theoretical study of the nervous system.
Cultural Neuroscience
Cultural neuroscience is the study of how cultural values, practices and beliefs shape and are shaped by the mind, brain and genes across multiple timescales.
Developmental Neuroscience
Developmental neuroscience studies the processes that generate, shape, and reshape the nervous system and seeks to describe the cellular basis of neural development to address underlying mechanisms.
Evolutionary Neuroscience
Evolutionary neuroscience studies the evolution of nervous systems.
Molecular Neuroscience
Molecular neuroscience studies the nervous system with molecular biology, molecular genetics, protein chemistry, and related methodologies.
Neural Neuroscience
Neural engineering uses engineering techniques to interact with, understand, repair, replace, or enhance neural systems.
Neuroanatomy
Neuroanatomy is the study of the anatomy of nervous systems.
Neurochemistry
Neurochemistry is the study of how neurochemicals interact and influence the function of neurons.
Neuroethology
Neuroethology is the study of the neural basis of non-human animals behaviour.
Neurogastronomy
Neurogastronomy is the study of flavour and how it affects sensation, cognition, and memory.
Neurogenetics
Neurogenetics is the study of the genetical basis of the development and function of the nervous system.
Neuroimaging
Neuroimaging includes the use of various techniques to either directly or indirectly image the structure and function of the brain.
Neuroimmunology
Neuroimmunology is concerned with the interactions between the nervous and the immune system.
Neuroinformatics
Neuroinformatics is a discipline within bioinformatics that conducts the organisation of neuroscience data and application of computational models and analytical tools.
Neurolinguistics
Neurolinguistics is the study of the neural mechanisms in the human brain that control the comprehension, production, and acquisition of language.
Neurophysics
Neurophysics deals with the development of physical experimental tools to gain information about the brain.
Neurophysiology
Neurophysiology is the study of the functioning of the nervous system, generally using physiological techniques that include measurement and stimulation with electrodes or optically with ion- or voltage-sensitive dyes or light-sensitive channels.
Neuropsychology
Neuropsychology is a discipline that resides under the umbrellas of both psychology and neuroscience, and is involved in activities in the arenas of both basic science and applied science. In psychology, it is most closely associated with biopsychology, clinical psychology, cognitive psychology, and developmental psychology. In neuroscience, it is most closely associated with the cognitive, behavioural, social, and affective neuroscience areas. In the applied and medical domain, it is related to neurology and psychiatry.
Paleoneurobiology
Paleoneurobiology is a field which combines techniques used in palaeontology and archaeology to study brain evolution, especially that of the human brain.
Social Neuroscience
Social neuroscience is an interdisciplinary field devoted to understanding how biological systems implement social processes and behaviour, and to using biological concepts and methods to inform and refine theories of social processes and behaviour.
Systems Neuroscience
Systems neuroscience is the study of the function of neural circuits and systems.
Neuroscience Organisations
The largest professional neuroscience organisation is the Society for Neuroscience (SFN), which is based in the United States but includes many members from other countries. Since its founding in 1969 the SFN has grown steadily: as of 2010 it recorded 40,290 members from 83 different countries. Annual meetings, held each year in a different American city, draw attendance from researchers, postdoctoral fellows, graduate students, and undergraduates, as well as educational institutions, funding agencies, publishers, and hundreds of businesses that supply products used in research.
Other major organisations devoted to neuroscience include the International Brain Research Organisation (IBRO), which holds its meetings in a country from a different part of the world each year, and the Federation of European Neuroscience Societies (FENS), which holds a meeting in a different European city every two years. FENS comprises a set of 32 national-level organisations, including the British Neuroscience Association, the German Neuroscience Society (Neurowissenschaftliche Gesellschaft), and the French Société des Neurosciences. The first National Honour Society in Neuroscience, Nu Rho Psi, was founded in 2006. Numerous youth neuroscience societies which support undergraduates, graduates and early career researchers also exist, like Project Encephalon.
In 2013, the BRAIN Initiative was announced in the US. An International Brain Initiative was created in 2017, currently integrated by more than seven national-level brain research initiatives (US, Europe, Allen Institute, Japan, China, Australia, Canada, Korea, Israel) spanning four continents.
Public Education and Outreach
In addition to conducting traditional research in laboratory settings, neuroscientists have also been involved in the promotion of awareness and knowledge about the nervous system among the general public and government officials. Such promotions have been done by both individual neuroscientists and large organisations. For example, individual neuroscientists have promoted neuroscience education among young students by organising the International Brain Bee, which is an academic competition for high school or secondary school students worldwide. In the United States, large organisations such as the Society for Neuroscience have promoted neuroscience education by developing a primer called Brain Facts, collaborating with public school teachers to develop Neuroscience Core Concepts for K-12 teachers and students, and cosponsoring a campaign with the Dana Foundation called Brain Awareness Week to increase public awareness about the progress and benefits of brain research. In Canada, the CIHR Canadian National Brain Bee is held annually at McMaster University.
Neuroscience educators formed Faculty for Undergraduate Neuroscience (FUN) in 1992 to share best practices and provide travel awards for undergraduates presenting at Society for Neuroscience meetings.
Finally, neuroscientists have also collaborated with other education experts to study and refine educational techniques to optimise learning among students, an emerging field called educational neuroscience. Federal agencies in the United States, such as the National Institute of Health (NIH) and National Science Foundation (NSF), have also funded research that pertains to best practices in teaching and learning of neuroscience concepts.
1930 – Dorothy Rowe, Australian psychologist and author (d. 2019).
1931 – James McGaugh, American neurobiologist and psychologist.
Dorothy Rowe
Dr. Dorothy Rowe (née Conn; 17 December 1930 to 25 March 2019) was an Australian psychologist and author, whose area of interest was depression. Born; Newcastle, NSW. Died Sydney, NSW.
Rowe came to England in her forties, working at Sheffield University and was the head of Lincolnshire Department of Clinical Psychology. In addition to her published works on depression, she is a regular columnist in the UK.
She spent her time working with depressed patients and, through listening to their stories, came to reject the medical model of mental illness, instead working within personal construct theory. She believes that depression is a result of beliefs which do not enable a person to live comfortably with themselves or the world. Most notably it is the belief in a “Just World” (that the bad are punished and the good rewarded) that exacerbates feelings of fear and anxiety if disaster strikes. Part of recovering is accepting that the external world is unpredictable and that we control relatively little of it.
In July 1989, Rowe made an extended appearance on the British television discussion programme After Dark alongside, among others, Steven Rose, Frank Cioffi, The Bishop of Durham and Michael Bentine.
The BBC were required to apologise to Dorothy Rowe in 2009 after the production editing of her radio interview misrepresented her views on the impact of religion in providing structure to people’s lives.
James McGaugh
James L. McGaugh (born 17 December 1931) is an American neurobiologist and author working in the field of learning and memory. He is a Distinguished Professor Emeritus in the Department of Neurobiology and Behaviour at the University of California, Irvine and a fellow and founding director of the Centre for the Neurobiology of Learning and Memory.
Career
McGaugh received his B.A. from San Jose State University in 1953 and his PhD in psychology from the University of California, Berkeley, in 1959. He was briefly a professor at San Jose State and then did postdoctoral work in neuropharmacology with Nobel Laureate Professor Daniel Bovet at the Istituto Superiore di Sanitá in Rome, Italy. He then became a professor at the University of Oregon from 1961 to 1964. He was recruited to the University of California, Irvine, in 1964 (the year of the school’s founding) to be the founding chair of the Department of Psychobiology (now Neurobiology and Behaviour). He became dean (1967-1970) of the School of Biological Sciences and Vice Chancellor (1975-1977) and executive Vice Chancellor (1978-1982) of the university. In 1982, he founded the Centre for the Neurobiology of Learning and Memory and remained director from 1982 to 2004.
Early Research Findings
McGaugh’s early work (in the 1950s and 1960s) demonstrated that memories are not instantly created in a long-term, permanent fashion. Rather, immediately after a learning event, the memory is labile and susceptible to influence. As time passes, the memory becomes increasingly resistant to external influences and eventually becomes stored in a relatively permanent manner, a process termed memory consolidation. McGaugh found that drugs, given to an animal shortly after a learning event, influence the subsequent retention of that event. The concept of such “post-training” manipulations is one of McGaugh’s greatest contributions to the field of learning and memory because it avoids many potential confounds, such as performance effects of the drug, that may occur when a drug or other treatment is given prior to the training.
Over the ensuing decades, McGaugh and his research colleagues and students extended the findings into a long-term investigation of emotionally influenced memory consolidation. As most people realise, they have stronger memories for long-ago events that were emotionally arousing in nature, compared with memories for emotionally neutral events (which may not be remembered well at all). McGaugh’s research examined how emotional arousal influences memory consolidation. In particular, he has found that stress hormones, such as epinephrine and cortisol, mediate much of the effects of emotional arousal on subsequent retention of the event. These hormones, in turn, activate a variety of brain structures, including the amygdala, which appears to play a key role in modulating memory consolidation. The amygdala, when activated, influences a variety of other brain structures, including the hippocampus, nucleus accumbens and caudate nucleus that process different aspects of memory. It is through this “orchestration” of brain structures that memories are eventually formed and stored, though the exact nature of memory storage remains elusive.
Author(s): Kathryn Eve Lewandowski and Ahmed A. Moustafa (Editors).
Year: 2019.
Edition: First (1st).
Publisher: Academic Press.
Type(s): Paperback and Kindle.
Synopsis:
Social Cognition in Psychosis combines current research on phenotypes, neurobiology, and existing evidence on the assessment and treatment of various forms of psychoses.
The book presents various treatment options, including assessment approaches, tools and training methods that aid in the rehabilitation of patients with psychotic disorders.
Social cognition is a set of psychological processes related to understanding, recognising, processing and appropriately using social stimuli in one’s environment.
Individuals with psychotic disorders consistently exhibit impairments in social cognition. As a result, social cognition has been an important target for intervention, with recent efforts trying to enhance early recovery among individuals with psychotic disorders.





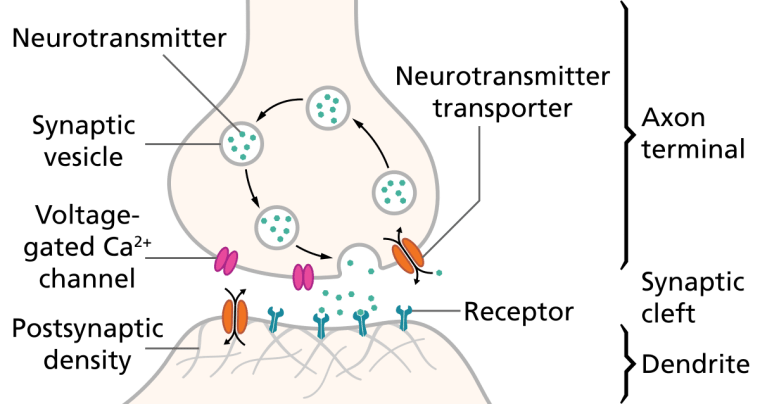
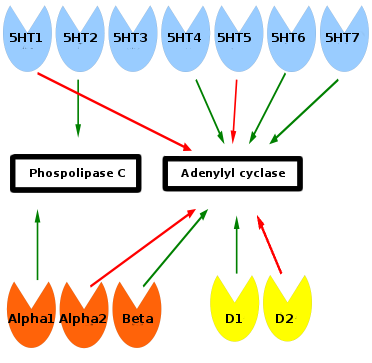
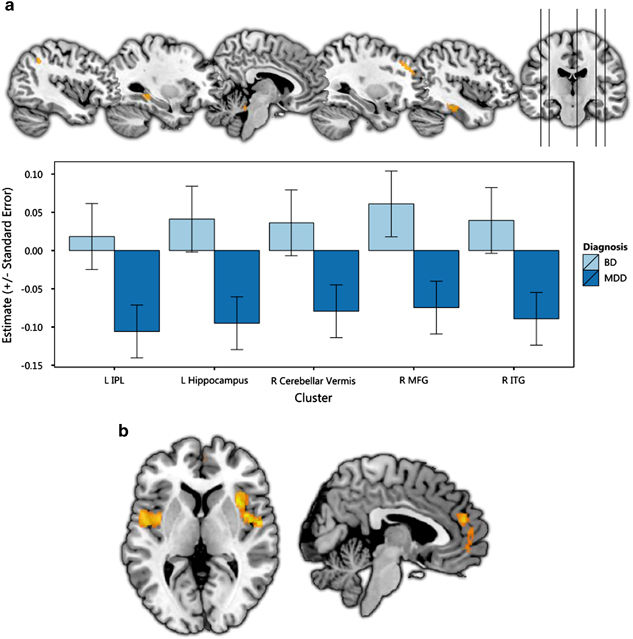





You must be logged in to post a comment.